За связь эмоций и формирования памяти отвечает гормон норэпинефрин
Новость
Графическая формула гормона норэпинефрина (норадреналина)
Результаты новых исследований на мышах добавляют ясности к вопросу о том, почему важные для нас моменты и крайне эмоциональные ситуации могут оставлять сильнейшие впечатления и держаться в памяти всю жизнь. Ученые из лаборатории в Cold Spring Harbor считают, что на эти процессы влияет гормон норэпинефрин (норадреналин).
«Этот феномен знаком всем и каждому, — говорит Роберто Малиноу (Roberto Malinow), один из авторов работы. — Вы с высокой вероятностью помните, где были в тот момент, когда узнали о случившемся 11 сентября, но при этом вы вряд ли можете сказать, что же вы делали днем ранее. Нам удалось идентифицировать один из механизмов, ответственных за подобный эффект» [1].
Исследователи объсняют, что отделы мозга, в которых хранятся воспоминания, должны как-то отличать важный и полезный опыт от ситуаций менее значимых, отдавая приоритет тем или иным впечатлениям при сохранении их в долговременной памяти.
Считается, что стрессовый гормон норэпинефрин играет одну из ключевых ролей в формировании связи между эмоциями и памятью. Во время эмоционального возбуждения этот гормон выделяется нейронами, связанными с большим количеством областей мозга, участвующих в формировании эмоциональной памяти.
Стимуляция мозга норэпинефрином, как было показано, индуцирует так называемую долговременную потенциацию (способность нейронов последовательно усиливать ответную реакцию на учащающиеся импульсы). Считается, что это свойство нейронов лежит в основе возможности обучения и формирования памяти.
До недавнего времени не был известен конкретный молекулярный механизм воздействия стрессовых гормонов на процессы формирование памяти. Однако группа Малиноу показала, что под воздействием норэпинефрина может увеличиваться количество мембранных рецепторов GluR1 в нейронах, от которого зависит эффективность работы синапса [2].
В исследовании на мышах ученые показали, что норэпинефрин, также как и эмоциональный стресс, провоцирует фосфорилирование молекул рецептора GluR1 по тем сайтам, которые необходимы для того, чтобы эти рецепторы локализовывались в постсинаптических мембранах. Такая модификация оказалась необходимой и достаточной для того, чтобы начала осуществляться долговременная потенциация, а следовательно, активизировались механизмы формирования памяти.
При последующем исследовании поведенческих реакций у подопытных животных, ученые обнаружили, что воздействие норэпинефрина позволяет мышам запоминать события с большей точностью. Тогда как мыши, мутантные по белкам GluR1 (особенно если мутации затрагивают фосфорилируемые домены), не демонстрировали улучшения памяти в ответ на норэпинефриновое воздействие.
По словам Малиноу, мозг мыши включает «все те же части», что и мозг человека, а тесты с исследованием связи памяти и эмоций у людей показываютс, что блокирование норэпинефриновых рецепторов снижает эффекты от воздействия сильных эмоций на процессы обучения и памяти. «Мы считаем, что молекулярные механизмы таких эффектов у людей не отличаются от того, что было продемонстрировано на мышах
», — говорит исследователь.Малиноу в тоже время обращает особое внимание на то, что настоящая работа — это всего лишь небольшой вклад в исследование того, как эмоции влияют на память. Также остается неясным, действительно ли обнаруженный механизм играет роль в формировании воспоминания при посттравматическом стрессе. Однако, как считают ученые, благодаря их результатам в будущем станет возможно создание препаратов, позволяющих регулировать количество рецепторов в нейронах мозга.
- How emotionally charged events leave their mark on memory. (2007). Biology News Net;
- Hailan Hu, Eleonore Real, Kogo Takamiya, Myoung-Goo Kang, Joseph Ledoux, et.
 al.. (2007). Emotion Enhances Learning via Norepinephrine Regulation of AMPA-Receptor Trafficking. Cell. 131, 160-173.
al.. (2007). Emotion Enhances Learning via Norepinephrine Regulation of AMPA-Receptor Trafficking. Cell. 131, 160-173.
 al.. (2007). Emotion Enhances Learning via Norepinephrine Regulation of AMPA-Receptor Trafficking. Cell. 131, 160-173.
al.. (2007). Emotion Enhances Learning via Norepinephrine Regulation of AMPA-Receptor Trafficking. Cell. 131, 160-173.Память: способность мозга хранить и восстанавливать информацию
Что такое Память?
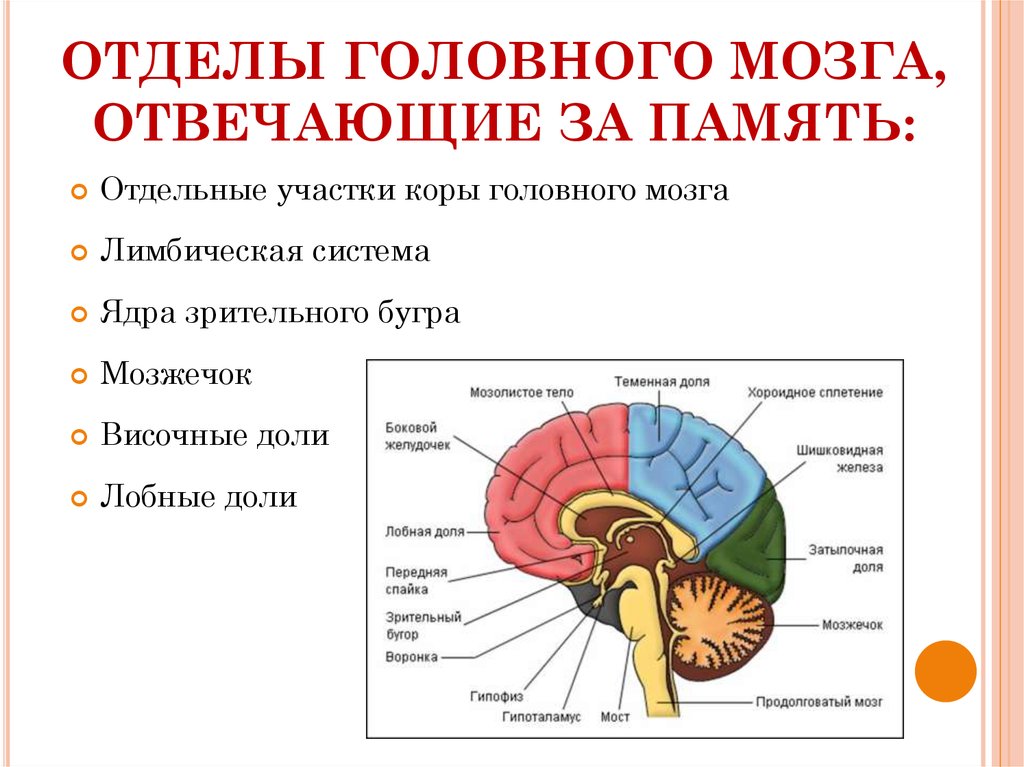
Память можно определить как способность мозга удерживать и добровольно восстанавливать информацию. Другими словами, это способность, которая позволяет нам вспоминать произошедшие события, мысли, ощущения, понятия и взаимосвязь между ними. Несмотря на то, что больше всего с памятью связан гиппокамп, отнести воспоминания только к одному отделу мозга нельзя, поскольку в этом процессе задействованы множество областей нашего мозга. Эта способность является одной из когнитивных функций, наиболее страдающих при старении. К счастью, память можно тренировать с помощью когнитивной стимуляции и различных умных игр.
Программа CogniFit («КогниФит»), являющаяся лидером в области тренировки мозга, позволяет укрепить эту и другие важнейшие когнитивные способности. Входящие в программу умные игры были разработаны для стимулирования определённых нейронных паттернов активации. Повторение этих когнитивных паттернов помогает укрепить задействованные в памяти нейронные связи, а также содействует созданию новых синапсов, способных реорганизовать и/или восстановить наиболее ослабленные или пострадавшие когнитивные функции.
Входящие в программу умные игры были разработаны для стимулирования определённых нейронных паттернов активации. Повторение этих когнитивных паттернов помогает укрепить задействованные в памяти нейронные связи, а также содействует созданию новых синапсов, способных реорганизовать и/или восстановить наиболее ослабленные или пострадавшие когнитивные функции.
Виды памяти
Память — чрезвычайно сложная когнитивная функция. В ней участвует огромное количество отделов мозга, и мы постоянно её используем. Существуют различные теории и исследования этой когнитивной способности. Можно подразделить память на различные виды по следующим критериям:
- По времени, в течение которого удерживается информация: в данном случае речь идёт о сенсорной памяти, кратковременной памяти, рабочей памяти и долговременной памяти. Сенсорная память удерживает информацию в течение нескольких секунд, в то время как долговременная память, наоборот, может хранить информацию в течение практически неограниченного периода времени. Все виды памяти работают скоординированно для того, чтобы вся система функционировала корректно.
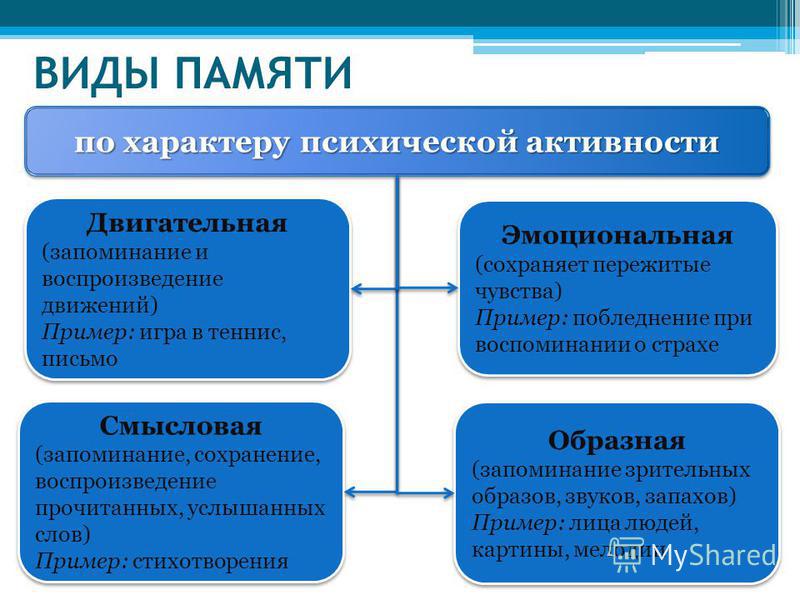
- По типу информации: вербальная память отвечает за хранение вербальной информации (то, что мы читаем, или слова, которые мы слышим), в то время как невербальная память позволяет хранить остальные данные (изображения, звуки, ощущения и т.д.).
- По задействованному органу чувств: в зависимости от используемого органа чувств, речь идёт о таких видах памяти, как зрительная память (зрение), слуховая память (слух), обонятельная память (обоняние), вкусовая память (вкус) и тактильная память (осязание).
 Все виды памяти работают скоординированно для того, чтобы вся система функционировала корректно.
Все виды памяти работают скоординированно для того, чтобы вся система функционировала корректно.Фазы памяти: процесс запоминания и воспоминания
Для того, чтобы вспомнить, что мы делали вчера, наш мозг должен произвести серию процессов. Каждый процесс необходим для доступа к воспоминаниям. Таким образом, нарушение любого из этих процессов не позволит нам вспомнить информацию. Чтобы создать воспоминание, наш мозг должен пройти через следующие фазы:
- Кодирование: на этой фазе в нашу систему памяти с помощью восприятия мы добавляем информацию, которую мы запоминаем. Например, когда нам кого-то представляют по имени. Необходимо обратить внимание на эту информацию, чтобы закодировать её.
- Хранение: чтобы удержать информацию надолго, мы сохраняем её в системе нашей памяти. Например, мы можем запомнить лицо человека и его имя.
- Восстановление: когда мы хотим что-то вспомнить, то обращаемся к хранилищу памяти и восстанавливаем нужную информацию. Например, чтобы, увидев человека на улице, вспомнить, как его зовут.
 Например, когда нам кого-то представляют по имени. Необходимо обратить внимание на эту информацию, чтобы закодировать её.
Например, когда нам кого-то представляют по имени. Необходимо обратить внимание на эту информацию, чтобы закодировать её.Примеры памяти
- Благодаря этой способности мы помним, где живём, как зовут наших родителей, лица наших друзей, что мы ели вчера на обед и даже какой город является столицей нашего государства.
- Память позволяет нам вспомнить о собрании на работе, запомнить имя клиента или пароль на компьютере.
- Учиться в школе или университете было бы невозможно без нашей системы хранения воспоминаний. Также нам было бы сложно запомнить дату экзамена или что мы запланировали сделать.
- При вождении автомобиля данная способность помогает нам вспомнить нужный маршрут. Также с её помощью мы помним, где припарковали машину, да и сам процесс вождения.

Амнезия и другие расстройства памяти
Исследования нарушений данной когнитивной функции помогли выяснить, что на самом деле представляет из себя память и как она работает. Являясь сложнейшей когнитивной функцией, она может пострадать в разной степени и по разным причинам. С одной стороны, специфические поражения могут быть связаны с двойной диссоциацией систем памяти. Это означает, что может быть повреждена одна из систем, в то время как другие не пострадают (например, может быть нарушена долговременная память при нормальном функционировании кратковременной). С другой стороны, подобные расстройства могут быть связаны с нейродегенеративным заболеванием (деменции и болезнь Альцгеймера), приобретённым церебральным поражением (черепно-мозговые травмы, инсульт, инфекции и другие болезни), врождёнными проблемами (паралич мозга и другие синдромы), с психическими расстройствами и расстройствами настроения (шизофрения, депрессия и тревожность), потреблением различных веществ (наркотики и медикаменты) и т. д. Также отдельные виды памяти могут быть нарушены при таких расстройствах обучаемости, как СДВГ, дислексия или дискалькулия.
д. Также отдельные виды памяти могут быть нарушены при таких расстройствах обучаемости, как СДВГ, дислексия или дискалькулия.
Наиболее распространённым расстройством памяти является потеря памяти, например, при болезни Альцгеймера. Потеря этой способности известна как амнезия. Амнезии бывают антероградные (неспособность приобрести новые воспоминания) и ретроградные (неспособность вспомнить прошлое). Также существуют расстройства, при которых нарушено содержание воспоминаний (фабуляции и конфабуляции), гипермнезии. Характерные для Синдрома Корсакова конфабуляции представляют собой непроизвольные ложные воспоминания, при которых забытая информация заменяется вымышленными фактами. Гипермнезии, в свою очередь, представляют собой непроизвольное и слишком детальное воспоминание малозначимых, несущественных деталей, что характерно, в частности, при посттравматическом стрессе.
Как можно измерить и оценить состояние нашей памяти?
Тестирование состояния нашей памяти очень полезно, поскольку она имеет важнейшее значение в учебной сфере (для того, чтобы понимать, будет ли ребёнок испытывать трудности с запоминанием пройденного материала и нуждается ли в дополнительной помощи), в медицине (для того, чтобы понимать, будет ли помнить пациент какие ему нужно принимать лекарства, может ли он быть самостоятельным или ему необходима помощь), на работе (для того, чтобы понимать, может ли человек занимать определённую должность) и в нашей повседневной жизни
С помощью комплексного нейропсихологического тестирования можно надёжно и эффективно измерить память и другие когнитивные функции. CogniFit («КогниФит») предлагает серию тестов, которые оценивают некоторые субпроцессы памяти, такие как кратковременная фонологическая память, контекстуальная память, кратковременная память, невербальная память, кратковременная зрительная память, рабочая память и распознавание. Тесты CogniFit («КогниФит») основаны на классическом Тесте на Длительное Поддержание Функции (CPT, Тест Коннера), Шкале Памяти Векслера (WMS), NEPSY (Коркман, Кирк и Кемп), Тесте Переменных Внимания (TOVA), Тесте на Симуляцию Нарушений Памяти (TOMM), Тесте «Лондонская башня» (TOL) и Задаче Визуальной Организации Хупера (VOT). С помощью этих тестов кроме памяти также можно измерить время отклика или реакции, скорость обработки информации, память на имена, зрительное восприятие, мониторинг, планирование, визуальное сканирование и пространственное восприятие.
CogniFit («КогниФит») предлагает серию тестов, которые оценивают некоторые субпроцессы памяти, такие как кратковременная фонологическая память, контекстуальная память, кратковременная память, невербальная память, кратковременная зрительная память, рабочая память и распознавание. Тесты CogniFit («КогниФит») основаны на классическом Тесте на Длительное Поддержание Функции (CPT, Тест Коннера), Шкале Памяти Векслера (WMS), NEPSY (Коркман, Кирк и Кемп), Тесте Переменных Внимания (TOVA), Тесте на Симуляцию Нарушений Памяти (TOMM), Тесте «Лондонская башня» (TOL) и Задаче Визуальной Организации Хупера (VOT). С помощью этих тестов кроме памяти также можно измерить время отклика или реакции, скорость обработки информации, память на имена, зрительное восприятие, мониторинг, планирование, визуальное сканирование и пространственное восприятие.
- Последовательный Тест WOM-ASM: на экране появится серия шаров с различными цифрами. Необходимо запомнить эту серию цифр, чтобы затем воспроизвести её. Сначала серия будет состоять только из одной цифры, затем количество цифр будет расти до тех пор, пока пользователь не совершит ошибку. Нужно будет повторить каждую представленную серию.
- Тест-Расследование REST-COM: в течение короткого промежутка времени будут представлены объекты. Далее как можно быстрее нужно будет выбрать слово, соответствующее показанному изображению.
- Тест Идентификации COM-NAM: объекты будут представлены с помощью изображения или звука. Необходимо ответить в каком формате объект был показан в последний раз и был ли показан вообще.
- Тест на Концентрацию VISMEM-PLAN: на экране в случайном порядке появятся стимулы. Стимулы начнут загораться в определённой последовательности под звуковые сигналы. Необходимо обратить внимание как на звуки, так и на последовательность световых сигналов. Во время очереди игры пользователя нужно воспроизвести увиденный ранее порядок представления стимулов.
- Тест на Распознавание WOM-REST: на экране появятся три объекта. Сначала нужно будет как можно быстрее вспомнить порядок представления этих объектов. Далее появятся четыре серии по три объекта, некоторые из которых будут отличаться от ранее увиденных. Необходимо восстановить первоначальную последовательность в том же порядке.
- Тест на Восстановление VISMEM: в течение пяти-шести секунд на экране будет представлено изображение. За это время нужно постараться запомнить максимальное количество объектов на этом изображении. Затем картинка исчезнет, и пользователь должен будет выбрать верный вариант ответа из предложенных.
 Сначала серия будет состоять только из одной цифры, затем количество цифр будет расти до тех пор, пока пользователь не совершит ошибку. Нужно будет повторить каждую представленную серию.
Сначала серия будет состоять только из одной цифры, затем количество цифр будет расти до тех пор, пока пользователь не совершит ошибку. Нужно будет повторить каждую представленную серию. Сначала нужно будет как можно быстрее вспомнить порядок представления этих объектов. Далее появятся четыре серии по три объекта, некоторые из которых будут отличаться от ранее увиденных. Необходимо восстановить первоначальную последовательность в том же порядке.
Сначала нужно будет как можно быстрее вспомнить порядок представления этих объектов. Далее появятся четыре серии по три объекта, некоторые из которых будут отличаться от ранее увиденных. Необходимо восстановить первоначальную последовательность в том же порядке.Восстановить, улучшить и стимулировать память
Все когнитивные способности, включая память, можно улучшить с помощью тренировки. CogniFit («КогниФит») даёт возможность делать это профессионально.
Пластичность мозга является основой для реабилитации памяти и других когнитивных функций. Мозг и его нейронные связи укрепляются за счёт использования функций, которые от них зависят. Таким образом, при тренировке памяти укрепляются нейронные связи задействованных отделов мозга.
Таким образом, при тренировке памяти укрепляются нейронные связи задействованных отделов мозга.
CogniFit («КогниФит») состоит из опытной команды профессионалов, специализирующихся на изучении синаптической пластичности и процессов нейрогенеза. Это позволило создать персонализированную программу когнитивной стимуляции для каждого пользователя. Программа начинается с точной оценки памяти и других основных когнитивных функций. По итогам тестирования программа когнитивной стимуляции Cognifit («КогниФит») автоматически предложит персональную когнитивную тренировку для улучшения памяти и других когнитивных функций, которые, согласно оценке, в этом нуждаются.
Чтобы улучшить память, тренироваться нужно правильно и регулярно. CogniFit («КогниФит») предлагает инструменты оценки и реабилитации памяти и других когнитивных функций. Для корректной стимуляции необходимо 15 минут в день, два или три раза в неделю.
Эта программа доступна онлайн. Разнообразные интерактивные упражнения представлены в виде увлекательных умных игр, в которые можно играть с помощью компьютера. В конце каждой сессии CogniFit («КогниФит») представит подробный график прогресса когнитивного состояния.
Нейробиологические основы формирования памяти: от физиологических состояний к психопатологии
1. Кандель Э.Р. Молекулярная биология хранения памяти: диалог между генами и синапсами. Наука. 2001; 294:1030–1038. [PubMed] [Google Scholar]
2. Дудай Ю. Нейробиология консолидаций, или насколько стабильна инграмма? Ежегодный обзор психологии. 2004; 55: 51–86. [PubMed] [Google Scholar]
3. Lamprecht R, Farb CR, Rodrigues SM, LeDoux JE. Обусловливание страха направляет профилин в дендритные шипы миндалевидного тела. Нат Нейроски. 2006;9: 481–483. [PubMed] [Google Scholar]
4. Альберини CM. Роль синтеза белка во время лабильных фаз памяти: пересмотр скептицизма. Нейробиол Узнать Мем. 2008; 89: 234–246. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Нейробиол Узнать Мем. 2008; 89: 234–246. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Alberini CM. Факторы транскрипции в долговременной памяти и синаптической пластичности. Physiol Rev. 2009; 89: 121–145. [Бесплатная статья PMC] [PubMed] [Google Scholar]
6. Davis HP, Squire LR. Синтез белка и память: обзор. Психологический бюллетень. 1984;96:518–559. [PubMed] [Google Scholar]
7. Граф П., Шактер Д.Л. Имплицитная и эксплицитная память на новые ассоциации у нормальных и страдающих амнезией субъектов. Журнал экспериментальной психологии Обучение, память и познание. 1985; 11: 501–518. [PubMed] [Google Scholar]
8. Squire LR, Zola SM. Структура и функции декларативной и недекларативной систем памяти. Proc Natl Acad Sci U S A. 1996; 93:13515–13522. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Карони П., Донато Ф., Мюллер Д. Структурная пластичность при обучении: регуляция и функции. Нат Рев Нейроски. 2012; 13: 478–49.0. [PubMed] [Google Scholar]
10. Frankland PW, Bontempi B. Организация недавних и отдаленных воспоминаний. Нат Рев Нейроски. 2005; 6: 119–130. [PubMed] [Google Scholar]
Frankland PW, Bontempi B. Организация недавних и отдаленных воспоминаний. Нат Рев Нейроски. 2005; 6: 119–130. [PubMed] [Google Scholar]
11. Дудай Ю. Беспокойная инграмма: Консолидации никогда не заканчиваются. Ежегодный обзор неврологии. 2012; 35: 227–247. [PubMed] [Google Scholar]
12. Рамамурти Б. Четвертое состояние сознания: турия авастха. Психиатрия и клиническая неврология. 1995; 49: 107–110. [PubMed] [Академия Google]
13. Дудай Ю., Айзенберг М. Ритуалы прохождения инграммы: реконсолидация и гипотеза затяжной консолидации. Нейрон. 2004; 44: 93–100. [PubMed] [Google Scholar]
14. McGaugh JL. Память — столетие консолидации. Наука. 2000; 287: 248–251. [PubMed] [Google Scholar]
15. Alberini CM, Ledoux JE. Реконсолидация памяти. Карр Биол. 2013; 23: Р746–750. [PubMed] [Google Scholar]
16. Надер К., Эйнарссон Э.О. Реконсолидация памяти: обновление. Энн Н.Ю. Академия наук. 2010;1191:27–41. [PubMed] [Google Scholar]
17. Tronson NC, Taylor JR. Молекулярные механизмы реконсолидации памяти. Нат Рев Нейроски. 2007; 8: 262–275. [PubMed] [Google Scholar]
Молекулярные механизмы реконсолидации памяти. Нат Рев Нейроски. 2007; 8: 262–275. [PubMed] [Google Scholar]
18. Сара SJ. Поиск и реконсолидация: к нейробиологии запоминания. Выучить Мем. 2000;7:73–84. [PubMed] [Google Scholar]
19. Альберини CM. Роль реконсолидации и динамического процесса формирования и хранения долговременной памяти. Фронт Behav Neurosci. 2011;5:12. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Питман РК. Предложит ли блокада реконсолидации новое лечение посттравматического стрессового расстройства? Фронт Behav Neurosci. 2011;5:11. [Бесплатная статья PMC] [PubMed] [Google Scholar]
21. Hartley CA, Phelps EA. Изменение страха: нейросхема регуляции эмоций. Нейропсихофармакол. 2010; 35: 136–146. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Schafe GE, Nader K, Blair HT, LeDoux JE. Консолидация памяти павловского кондиционирования страха: клеточная и молекулярная перспектива. Тренды Нейроси. 2001; 24: 540–546. [PubMed] [Академия Google]
23. Сквайр Л.Р., Викстед Дж.Т. Когнитивная нейронаука человеческой памяти с тех пор, как Х.М. Ежегодный обзор неврологии. 2011; 34: 259–288. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Сквайр Л.Р., Викстед Дж.Т. Когнитивная нейронаука человеческой памяти с тех пор, как Х.М. Ежегодный обзор неврологии. 2011; 34: 259–288. [Бесплатная статья PMC] [PubMed] [Google Scholar]
24. Аргуэлло А.А., Йе Х., Боздаги О., Поллонини Г., Тронель С., Бамбах-Мукку Д., Хантли Г.В., Платано Д., Альберини К.М. Дельта белка, связывающего энхансер Ccaat, играет существенную роль в консолидации и реконсолидации памяти. Дж. Нейроски. 2013; 33:3646–3658. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Milekic MH, Pollonini G, Alberini CM. Временная потребность c/ebpbeta в миндалевидном теле после реактивации, но не приобретения ингибиторного избегания. Выучить Мем. 2007; 14: 504–511. [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Таубенфельд С.М., Милекич М.Х., Монти Б., Альберини К.М. Для консолидации новой, но не реактивированной памяти требуется c/ebpbeta гиппокампа. Нат Нейроски. 2001; 4: 813–818. [PubMed] [Google Scholar]
27. Кандель Э.Р. Молекулярная биология памяти: Camp, pka, cre, creb-1, creb-2 и cpeb. Молекулярный мозг. 2012;5:14. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Молекулярный мозг. 2012;5:14. [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Alberini CM, Chen DY. Улучшение памяти: консолидация, реконсолидация и инсулиноподобный фактор роста 2. Trends Neurosci. 2012 [Бесплатная статья PMC] [PubMed] [Google Scholar]
29. Стерн С.А., Альберини С.М. Механизмы улучшения памяти. Междисциплинарные обзоры Wiley по системной биологии и медицине. 2013; 5:37–53. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. de Kloet ER, Joels M, Holsboer F. Стресс и мозг: от адаптации к болезни. Нат Рев Нейроски. 2005; 6: 463–475. [PubMed] [Google Scholar]
31. de Quervain DJ, Aerni A, Schelling G, Roozendaal B. Глюкокортикоиды и регуляция памяти в норме и при болезни. Передний нейроэндокринол. 2009 г.;30:358–370. [PubMed] [Google Scholar]
32. Сэнди С., Рихтер-Левин Г. От черты высокой тревожности к депрессии: нейрокогнитивная гипотеза. Тренды Нейроси. 2009; 32: 312–320. [PubMed] [Google Scholar]
33. Питман Р. К., Расмуссон А.М., Коенен К.С., Шин Л.М., Орр С.П., Гилбертсон М.В., Милад М.Р., Либерзон И. Биологические исследования посттравматического стрессового расстройства. Нат Рев Нейроски. 2012;13:769–787. [Бесплатная статья PMC] [PubMed] [Google Scholar]
К., Расмуссон А.М., Коенен К.С., Шин Л.М., Орр С.П., Гилбертсон М.В., Милад М.Р., Либерзон И. Биологические исследования посттравматического стрессового расстройства. Нат Рев Нейроски. 2012;13:769–787. [Бесплатная статья PMC] [PubMed] [Google Scholar]
34. McEwen BS, Gianaros PJ. Пластичность мозга, вызванная стрессом и аллостазом. Ежегодный обзор медицины. 2011;62:431–445. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Кооб Г., Крик М.Дж. Стресс, нарушение регуляции путей вознаграждения за наркотики и переход к наркотической зависимости. Am J Психиатрия. 2007; 164:1149–1159. [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Roozendaal B, McGaugh JL. Модуляция памяти. Поведение Нейроски. 2011; 125:797–824. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Okuda S, Roozendaal B, McGaugh JL. Влияние глюкокортикоидов на память распознавания объектов требует связанного с обучением эмоционального возбуждения. Proc Natl Acad Sci U S A. 2004; 101:853–858. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
38. Санди С, Пинело-Нава, МТ. Стресс и память: поведенческие эффекты и нейробиологические механизмы. Нейр Пласт. 2007; 2007:78970. [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Revest JM, Di Blasi F, Kitchener P, Rouge-Pont F, Desmedt A, Turiault M, Tronche F, Piazza PV. Путь mapk и egr-1 опосредуют связанные со стрессом поведенческие эффекты глюкокортикоидов. Нат Нейроски. 2005; 8: 664–672. [PubMed] [Google Scholar]
40. Revest JM, Kaouane N, Mondin M, Le Roux A, Rouge-Pont F, Vallee M, Barik J, Tronche F, Desmedt A, Piazza PV. Улучшение связанной со стрессом памяти глюкокортикоидами зависит от синапсин-ia/ib. Мол Психиатрия. 2010;15:1125, 1140–1151. [Бесплатная статья PMC] [PubMed] [Google Scholar]
41. Чен Д.Ю., Бамбах-Мукку Д., Поллонини Г., Альберини К.М. Глюкокортикоидные рецепторы задействуют пути camkiialpha-bdnf-creb для обеспечения консолидации памяти. Нат Нейроски. 2012;15:1707–1714. [Бесплатная статья PMC] [PubMed] [Google Scholar]
42. Bilang-Bleuel A, Ulbricht S, Chandramohan Y, De Carli S, Droste SK, Reul JM. Психологический стресс увеличивает фосфорилирование гистона h4 в нейронах гранул зубчатой извилины взрослых: участие в поведенческом ответе, зависящем от глюкокортикоидных рецепторов. Евр Джей Нейроски. 2005;22:1691–1700. [PubMed] [Google Scholar]
Bilang-Bleuel A, Ulbricht S, Chandramohan Y, De Carli S, Droste SK, Reul JM. Психологический стресс увеличивает фосфорилирование гистона h4 в нейронах гранул зубчатой извилины взрослых: участие в поведенческом ответе, зависящем от глюкокортикоидных рецепторов. Евр Джей Нейроски. 2005;22:1691–1700. [PubMed] [Google Scholar]
43. Чандрамохан Ю., Дросте С.К., Артур Дж.С., Реул Дж.М. Реакция поведенческой неподвижности, вызванная вынужденным плаванием, включает фосфоацетилирование гистона h4 и индукцию c-fos в нейронах гранул зубчатой извилины посредством активации n-метил-d-аспартатной/внеклеточной сигнал-регулируемой киназы/митоген- и стресс-активируемой киназы. путь. Евр Джей Нейроски. 2008; 27: 2701–2713. [PubMed] [Google Scholar]
44. Розендаль Б., Эрнандес А., Кабрера С.М., Хагевуд Р., Мальваез М., Стефанко Д.П., Хеттиг Дж., Вуд М.А. Связанная с мембраной глюкокортикоидная активность необходима для модуляции долговременной памяти посредством модификации хроматина. Дж. Нейроски. 2010;30:5037–5046. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Дж. Нейроски. 2010;30:5037–5046. [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Финстервальд С., Альберини С.М. Стресс и механизмы, зависящие от глюкокортикоидных рецепторов, в долговременной памяти: от адаптивных реакций до психопатологий. Нейробиол Узнать Мем. 2013 [бесплатная статья PMC] [PubMed] [Google Scholar]
46. Allen SJ, Dawbarn D. Клиническая значимость нейротрофинов и их рецепторов. Clin Sci (Лондон) 2006; 110: 175–191. [PubMed] [Google Scholar]
47. Думан Р.С., Хенингер Г.Р., Нестлер Э.Дж. Молекулярно-клеточная теория депрессии. Арх генерал психиатрия. 1997;54:597–606. [PubMed] [Google Scholar]
48. Думан Р.С., Монтеджиа Л.М. Нейротрофическая модель расстройств настроения, связанных со стрессом. Биол психиатрия. 2006;59:1116–1127. [PubMed] [Google Scholar]
49. Pariante CM, Lightman SL. Ось hpa при большой депрессии: классические теории и новые разработки. Тренды Нейроси. 2008; 31: 464–468. [PubMed] [Google Scholar]
50. Nemeroff CB, Vale WW. Нейробиология депрессии: пути к лечению и открытие новых лекарств. Дж. Клин Психиатрия. 2005; 66 (Приложение 7): 5–13. [PubMed] [Академия Google]
Nemeroff CB, Vale WW. Нейробиология депрессии: пути к лечению и открытие новых лекарств. Дж. Клин Психиатрия. 2005; 66 (Приложение 7): 5–13. [PubMed] [Академия Google]
51. Чен Б., Довлатшахи Д., МакКуин Г.М., Ван Д.Ф., Янг Л.Т. Повышенная иммунореактивность bdnf гиппокампа у субъектов, получавших антидепрессанты. Биол психиатрия. 2001; 50: 260–265. [PubMed] [Google Scholar]
52. Dwivedi Y, Rizavi HS, Conley RR, Roberts RC, Tamminga CA, Pandey GN. Измененная экспрессия гена нейротрофического фактора головного мозга и рецепторной тирозинкиназы b в посмертном мозге суицидентов. Архив общей психиатрии. 2003; 60: 804–815. [PubMed] [Академия Google]
53. Karege F, Vaudan G, Schwald M, Perroud N, La Harpe R. Уровни нейротрофинов в посмертном мозге жертв самоубийств и эффекты прижизненной диагностики и психотропных препаратов. Исследования мозга Молекулярные исследования мозга. 2005; 136: 29–37. [PubMed] [Google Scholar]
54. Karege F, Perret G, Bondolfi G, Schwald M, Bertschy G, Aubry JM. Снижение уровня нейротрофического фактора головного мозга в сыворотке крови у пациентов с тяжелой депрессией. Психиатрия рез. 2002; 109: 143–148. [PubMed] [Академия Google]
Снижение уровня нейротрофического фактора головного мозга в сыворотке крови у пациентов с тяжелой депрессией. Психиатрия рез. 2002; 109: 143–148. [PubMed] [Академия Google]
55. Парианте СМ. Глюкокортикоидный рецептор: часть решения или часть проблемы? Журнал психофармакологии. 2006; 20:79–84. [PubMed] [Google Scholar]
56. Nader K, Schafe GE, Le Doux JE. Воспоминания о страхе требуют синтеза белка в миндалевидном теле для реконсолидации после извлечения. Природа. 2000;406:722–726. [PubMed] [Google Scholar]
57. Альберини CM. Механизмы стабилизации памяти: консолидация и реконсолидация похожи или разные процессы? Тренды Нейроси. 2005; 28:51–56. [PubMed] [Академия Google]
58. Милекич М.Х., Альберини СМ. Временная потребность в синтезе белка после реактивации памяти. Нейрон. 2002; 36: 521–525. [PubMed] [Google Scholar]
59. Suzuki A, Josselyn SA, Frankland PW, Masushige S, Silva AJ, Kida S. Реконсолидация и угасание памяти имеют различные временные и биохимические признаки. Дж. Нейроски. 2004; 24:4787–4795. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Дж. Нейроски. 2004; 24:4787–4795. [Бесплатная статья PMC] [PubMed] [Google Scholar]
60. Frankland PW, Ding HK, Takahashi E, Suzuki A, Kida S, Silva AJ. Стабильность недавних и отдаленных контекстуальных воспоминаний о страхе. Выучить Мем. 2006; 13: 451–457. [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. Графф Дж., Джозеф Н.Ф., Хорн М.Е., Самиэй А., Мэн Дж., Сео Дж., Рей Д., Беро А.В., Фан Т.Х., Вагнер Ф., Холсон Э., Сюй Дж., Сунь Дж., Нив Р.Л., Мах Р.Х., Хаггарти SJ, Цай ЛХ. Эпигенетическое праймирование обновления памяти во время реконсолидации для ослабления отдаленных воспоминаний о страхе. Клетка. 2014; 156: 261–276. [Бесплатная статья PMC] [PubMed] [Google Scholar]
62. Debec J, LeDoux JE, Nader K. Клеточная и системная реконсолидация в гиппокампе. Нейрон. 2002; 36: 527–538. [PubMed] [Google Scholar]
63. Alberini CM, Johnson SA, Ye X. Механизмы и функции реконсолидации памяти. В: Альберини CM, редактор. Реконсолидация памяти. Том. 1. Академическая пресса; Сан-Диего: 2013. стр. 81–117. [Академия Google]
стр. 81–117. [Академия Google]
64. Tronel S, Milekic MH, Alberini CM. Связывание новой информации с реактивированной памятью требует механизмов консолидации, а не реконсолидации. PLoS биол. 2005;3:e293. [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Хупбах А., Гомес Р., Надель Л. Реконсолидация эпизодической памяти: обновление или путаница с источником? Память. 2009; 17: 502–510. [PubMed] [Google Scholar]
66. Хупбах А., Хардт О., Гомес Р., Надель Л. Динамика памяти: контекстно-зависимое обновление. Выучить Мем. 2008; 15: 574–579.. [PubMed] [Google Scholar]
67. Morris RG, Inglis J, Ainge JA, Olverman HJ, Tulloch J, Dudai Y, Kelly PA. Реконсолидация памяти: чувствительность пространственной памяти к ингибированию синтеза белка в дорсальном гиппокампе во время кодирования и поиска. Нейрон. 2006; 50: 479–489. [PubMed] [Google Scholar]
68. Forcato C, Argibay PF, Pedreira ME, Maldonado H. Реконсолидация человека не всегда происходит при восстановлении памяти: актуальность структуры напоминания. Нейробиол Узнать Мем. 2009 г.;91:50–57. [PubMed] [Google Scholar]
Нейробиол Узнать Мем. 2009 г.;91:50–57. [PubMed] [Google Scholar]
69. Forcato C, Rodriguez ML, Pedreira ME, Maldonado H. Реконсолидация у людей открывает декларативную память для поступления новой информации. Нейробиол Узнать Мем. 2010;93:77–84. [PubMed] [Google Scholar]
70. Taubenfeld SM, Riceberg JS, New AS, Alberini CM. Доклиническая оценка избирательного нарушения травматической памяти посредством постретривного ингибирования глюкокортикоидных рецепторов. Биол психиатрия. 2009; 65: 249–257. [Бесплатная статья PMC] [PubMed] [Google Scholar]
71. Голье Дж.А., Караманика К., Демария Р., Йехуда Р. Пилотное исследование мифепристона при посттравматическом синдроме, связанном с боевыми действиями. Исследования и лечение депрессии. 2012;2012:393251. [Бесплатная статья PMC] [PubMed] [Google Scholar]
72. de Quervain DJ, Henke K, Aerni A, Treyer V, McGaugh JL, Berthold T, Nitsch RM, Buck A, Roozendaal B, Hock C. Глюкокортикоид-индуцированный нарушение декларативной памяти связано со снижением кровотока в медиальных отделах височной доли. Евр Джей Нейроски. 2003; 17: 1296–1302. [PubMed] [Академия Google]
Евр Джей Нейроски. 2003; 17: 1296–1302. [PubMed] [Академия Google]
73. Sandi C. Глюкокортикоиды действуют на глутаматергические пути, влияя на процессы памяти. Тренды Нейроси. 2011; 34: 165–176. [PubMed] [Google Scholar]
74. Аэрни А., Трабер Р., Хок С., Рузендал Б., Шеллинг Г., Папасситиропулос А., Нич Р.М., Шнайдер У., де Кервен Д.Дж. Низкие дозы кортизола при симптомах посттравматического стрессового расстройства. Am J Психиатрия. 2004; 161:1488–1490. [PubMed] [Google Scholar]
75. Соравиа Л.М., Генрихс М., Эрни А., Марони С., Шеллинг Г., Элерт У., Рузендал Б., де Кервен Д.Дж. Глюкокортикоиды уменьшают фобический страх у людей. Proc Natl Acad Sci USA. 2006; 103:5585–559.0. [Бесплатная статья PMC] [PubMed] [Google Scholar]
76. de Quervain DJ, Bentz D, Michael T, Bolt OC, Wiederhold BK, Margraf J, Wilhelm FH. Глюкокортикоиды усиливают психотерапию, основанную на угашении. Proc Natl Acad Sci U S A. 2011;108:6621–6625. [Бесплатная статья PMC] [PubMed] [Google Scholar]
77. Hill MN, McEwen BS. Участие эндоканнабиноидной системы в нейроповеденческих эффектах стресса и глюкокортикоидов. Прог Нейропсихофармакол Биол Психиатрия. 2010; 34: 791–797. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Hill MN, McEwen BS. Участие эндоканнабиноидной системы в нейроповеденческих эффектах стресса и глюкокортикоидов. Прог Нейропсихофармакол Биол Психиатрия. 2010; 34: 791–797. [Бесплатная статья PMC] [PubMed] [Google Scholar]
78. Hill MN, Patel S, Campolongo P, Tasker JG, Wotjak CT, Bains JS. Функциональные взаимодействия между стрессом и эндоканнабиноидной системой: от синаптической сигнализации до поведенческого результата. Дж. Нейроски. 2010;30:14980–14986. [Бесплатная статья PMC] [PubMed] [Google Scholar]
79. Steiner MA, Wotjak CT. Роль эндоканнабиноидной системы в регуляции гипоталамо-гипофизарно-адренокортикальной оси. Прог Мозг Res. 2008; 170:397–432. [PubMed] [Google Scholar]
80. Морена М., Камполонго П. Эндоканнабиноидная система: эмоциональный буфер в модуляции функции памяти. Нейробиология обучения и памяти. 2014 [PubMed] [Академия Google]
81. Линь Х.К., Мао С.К., Гин П.В. Влияние инфузии агонистов рецептора cb1 внутрь миндалевидного тела на реконсолидацию испуга, вызванного вздрагиванием. Выучить Мем. 2006; 13: 316–321. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Выучить Мем. 2006; 13: 316–321. [Бесплатная статья PMC] [PubMed] [Google Scholar]
82. Campolongo P, Roozendaal B, Trezza V, Hauer D, Schelling G, McGaugh JL, Cuomo V. Эндоканнабиноиды в базолатеральной миндалине крысы улучшают консолидацию памяти и обеспечивают модуляцию глюкокортикоидов. памяти. Proc Natl Acad Sci U S A. 2009;106:4888–4893. [Бесплатная статья PMC] [PubMed] [Google Scholar]
83. Лавиолетт С.Р., Грейс А.А. Каннабиноиды усиливают пластичность эмоционального обучения в нейронах медиальной префронтальной коры через входы базолатеральной миндалевидного тела. Дж. Нейроски. 2006; 26: 6458–6468. [Бесплатная статья PMC] [PubMed] [Google Scholar]
84. Tan H, Lauzon NM, Bishop SF, Bechard MA, Laviolette SR. Интегрированная передача каннабиноидного рецептора cb1 в миндалевидно-префронтальном кортикальном пути модулирует пластичность нейронов и кодирование эмоциональной памяти. Кора головного мозга. 2010; 20:1486–149.6. [PubMed] [Google Scholar]
85. Tan H, Lauzon NM, Bishop SF, Chi N, Bechard M, Laviolette SR. Передача каннабиноидов в базолатеральной миндалевидном теле модулирует формирование памяти о страхе через функциональные входы в прелимбическую кору. Дж. Нейроски. 2011;31:5300–5312. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Tan H, Lauzon NM, Bishop SF, Chi N, Bechard M, Laviolette SR. Передача каннабиноидов в базолатеральной миндалевидном теле модулирует формирование памяти о страхе через функциональные входы в прелимбическую кору. Дж. Нейроски. 2011;31:5300–5312. [Бесплатная статья PMC] [PubMed] [Google Scholar]
86. Pamplona FA, Bitencourt RM, Takahashi RN. Краткосрочные и долгосрочные эффекты каннабиноидов на угасание контекстуальной памяти о страхе у крыс. Нейробиол Узнать Мем. 2008;90:290–293. [PubMed] [Google Scholar]
87. Памплона Ф.А., Предигер Р.Д., Пандольфо П., Такахаши Р.Н. Агонист каннабиноидных рецепторов win 55,212-2 способствует угасанию контекстуальной памяти о страхе и пространственной памяти у крыс. Психофармакология (Берл) 2006; 188: 641–649. [PubMed] [Google Scholar]
88. Чхатвал Дж.П., Дэвис М., Магущак К.А., Ресслер К.Дж. Усиление каннабиноидной нейротрансмиссии усиливает угасание условного страха. Нейропсихофармакол. 2005; 30: 516–524. [PubMed] [Академия Google]
89. Izquierdo A, Wellman CL, Holmes A. Кратковременный неконтролируемый стресс вызывает ретракцию дендритов в инфралимбической коре и устойчивость к исчезновению страха у мышей. Дж. Нейроски. 2006; 26: 5733–5738. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Izquierdo A, Wellman CL, Holmes A. Кратковременный неконтролируемый стресс вызывает ретракцию дендритов в инфралимбической коре и устойчивость к исчезновению страха у мышей. Дж. Нейроски. 2006; 26: 5733–5738. [Бесплатная статья PMC] [PubMed] [Google Scholar]
90. Rabinak CA, Angstadt M, Sripada CS, Abelson JL, Liberzon I, Milad MR, Phan KL. Каннабиноидное облегчение воспоминаний об исчезновении страха у людей. Нейрофармакология. 2013; 64: 396–402. [Бесплатная статья PMC] [PubMed] [Google Scholar]
91. Izquierdo I, Quillfeldt JA, Zanatta MS, Quevedo J, Schaeffer E, Schmitz PK, Medina JH. Последовательная роль гиппокампа и миндалевидного тела, энторинальной коры и теменной коры в формировании и восстановлении памяти для торможения избегания у крыс. Евр Джей Нейроски. 1997; 9: 786–793. [PubMed] [Google Scholar]
92. Лутц Б. Эндоканнабиноидная система и обучение вымиранию. Мол Нейробиол. 2007; 36: 92–101. [PubMed] [Google Scholar]
93. Hill MN, Gorzalka BB. Эндоканнабиноидная система и лечение аффективных и тревожных расстройств. Лекарственные мишени для нейролептиков ЦНС. 2009 г.;8:451–458. [PubMed] [Google Scholar]
Лекарственные мишени для нейролептиков ЦНС. 2009 г.;8:451–458. [PubMed] [Google Scholar]
94. Trezza V, Campolongo P. Эндоканнабиноидная система как возможная мишень для лечения как когнитивных, так и эмоциональных особенностей посттравматического стрессового расстройства (ПТСР). Фронт Behav Neurosci. 2013;7:100. [Бесплатная статья PMC] [PubMed] [Google Scholar]
95. Monfils MH, Cowansage KK, Klann E, LeDoux JE. Границы угасания-реконсолидации: ключ к стойкому ослаблению воспоминаний о страхе. Наука. 2009; 324:951–955. [Бесплатная статья PMC] [PubMed] [Google Scholar]
96. Schiller D, Monfils MH, Raio CM, Johnson DC, Ledoux JE, Phelps EA. Предотвращение возвращения страха у людей с помощью механизмов обновления реконсолидации. Природа. 2010; 463:49–53. [Бесплатная статья PMC] [PubMed] [Google Scholar]
97. Jones CE, Monfils MH. Использование реконсолидации и угасания для ослабления воспоминаний о страхе на животных моделях. В: Альберини CM, редактор. Реконсолидация памяти.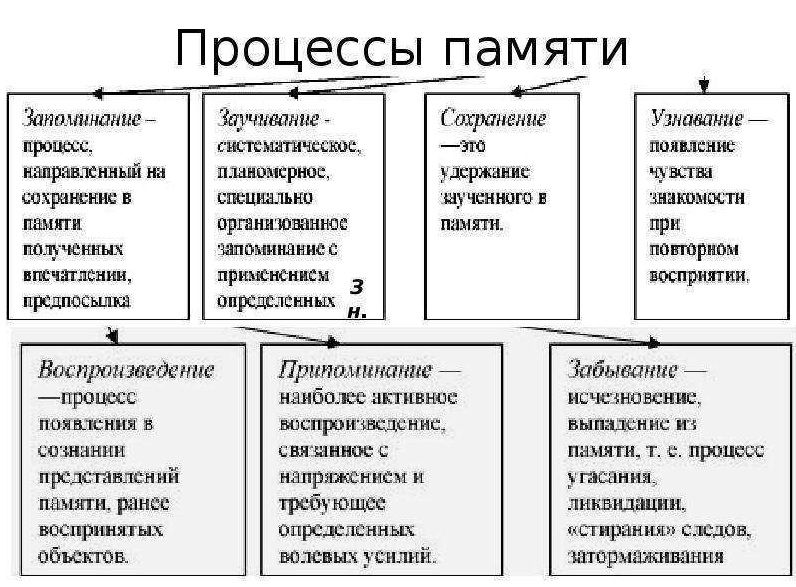 Том. 1. Академическая пресса; Сан-Диего: 2013. стр. 165–184. [Google Scholar]
Том. 1. Академическая пресса; Сан-Диего: 2013. стр. 165–184. [Google Scholar]
98. Фелпс Э.А., Шиллер Д. Реконсолидация у человека. В: Альберини CM, редактор. Реконсолидация памяти. Том. 1. Академическая пресса; Сан-Диего: 2013. стр. 185–212. [Академия Google]
99. Xue YX, Luo YX, Wu P, Shi HS, Xue LF, Chen C, Zhu WL, Ding ZB, Bao YP, Shi J, Epstein DH, Shaham Y, Lu L. Процедура извлечения-угасания памяти для предотвратить тягу к наркотикам и рецидивы. Наука. 2012; 336: 241–245. [Бесплатная статья PMC] [PubMed] [Google Scholar]
100. van Oostrom I, Dollfus S, Brazo P, Abadie P, Halbecq I, Thery S, Marie RM. Вербальное обучение и память у больных шизофренией и болезнью Паркинсона. Психиатрия рез. 2003; 117: 25–34. [PubMed] [Академия Google]
101. Чирилло М.А., Сейдман Л.Дж. Вербальная декларативная дисфункция памяти при шизофрении: от клинической оценки до генетики и механизмов мозга. Обзор нейропсихологии. 2003; 13:43–77. [PubMed] [Google Scholar]
102. Тулопулуанд Т., Мюррей Р.М. Дефицит вербальной памяти у пациентов с шизофренией: важная будущая цель лечения. Эксперт преподобный Нейротер. 2004; 4:43–52. [PubMed] [Google Scholar]
Тулопулуанд Т., Мюррей Р.М. Дефицит вербальной памяти у пациентов с шизофренией: важная будущая цель лечения. Эксперт преподобный Нейротер. 2004; 4:43–52. [PubMed] [Google Scholar]
103. Стефан К.Е., Балдевег Т., Фристон К.Дж. Синаптическая пластичность и разъединение при шизофрении. Биол психиатрия. 2006; 59: 929–939. [PubMed] [Google Scholar]
104. Favalli G, Li J, Belmonte-de-Abreu P, Wong AH, Daskalakis ZJ. Роль bdnf в патофизиологии и лечении шизофрении. J Psychiatr Res. 2012; 46:1–11. [PubMed] [Google Scholar]
105. Ангелуччи Ф., Брене С., Мате А.А. Bdnf при шизофрении, депрессии и соответствующих животных моделях. Молекулярная психиатрия. 2005; 10: 345–352. [PubMed] [Google Scholar]
106. Muglia P, Vicente AM, Verga M, King N, Macciardi F, Kennedy JL. Ассоциация между геном bdnf и шизофренией. Молекулярная психиатрия. 2003; 8: 146–147. [PubMed] [Академия Google]
107. Отри А.Е., Монтеджиа Л.М. Нейротрофический фактор головного мозга и нервно-психические расстройства. Фармакологические обзоры. 2012; 64: 238–258. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Фармакологические обзоры. 2012; 64: 238–258. [Бесплатная статья PMC] [PubMed] [Google Scholar]
108. Вытилингам М., Хейм С., Ньюпорт Дж., Миллер А.Х., Андерсон Э., Бронен Р., Браммер М., Стаиб Л., Верметтен Э., Чарни Д.С., Немерофф С.Б., Бремнер Джей Ди. Детская травма, связанная с меньшим объемом гиппокампа у женщин с большой депрессией. Am J Психиатрия. 2002;159:2072–2080. [Бесплатная статья PMC] [PubMed] [Google Scholar]
109. Хэкман Д.А., Фарах М.Дж., Мини М.Дж. Социально-экономический статус и мозг: механистические выводы из исследований человека и животных. Нат Рев Нейроски. 2010; 11: 651–659. [Бесплатная статья PMC] [PubMed] [Google Scholar]
110. Pratchett LC, Yehuda R. Основы посттравматического стрессового расстройства: приводит ли ранняя жизненная травма к посттравматическому стрессовому расстройству у взрослых? Дев психопат. 2011; 23: 477–491. [PubMed] [Google Scholar]
111. Фергюссон Д.М., Лински М., Хорвуд Л.Дж. Подростковые результаты усыновления: 16-летнее продольное исследование. Журнал детской психологии и психиатрии и смежных дисциплин. 1995;36:597–615. [PubMed] [Google Scholar]
Журнал детской психологии и психиатрии и смежных дисциплин. 1995;36:597–615. [PubMed] [Google Scholar]
112. Беккет С., Моган Б., Раттер М., Касл Дж., Колверт Э., Грутьюз С., Креппнер Дж., Стивенс С., О’Коннор Т.Г., Сонуга-Барк Э.Дж. Сохраняются ли последствия ранней тяжелой депривации на когнитивные функции в раннем подростковом возрасте? Результаты исследования усыновленных детей из Англии и Румынии. Развитие ребенка. 2006; 77: 696–711. [PubMed] [Google Scholar]
113. Sanchez MM, Ladd CO, Plotsky PM. Ранний неблагоприятный опыт как фактор риска развития более поздней психопатологии: данные моделей грызунов и приматов. Дев психопат. 2001;13:419–449. [PubMed] [Google Scholar]
114. Heim C, Plotsky PM, Nemeroff CB. Важность изучения вклада раннего неблагоприятного опыта в нейробиологические данные о депрессии. Нейропсихофармакол. 2004; 29: 641–648. [PubMed] [Google Scholar]
115. Hanson M, Godfrey KM, Lillycrop KA, Burdge GC, Gluckman PD. Пластичность развития и происхождение неинфекционных заболеваний: теоретические соображения и эпигенетические механизмы. Прогресс в биофизике и молекулярной биологии. 2011; 106: 272–280. [PubMed] [Академия Google]
Прогресс в биофизике и молекулярной биологии. 2011; 106: 272–280. [PubMed] [Академия Google]
116. Чжан Т.И., Лабонте Б., Вэнь С.Л., Турецки Г., Мини М.Дж. Эпигенетические механизмы ранней регуляции окружающей средой экспрессии генов глюкокортикоидных рецепторов гиппокампа у грызунов и человека. Нейропсихофармакол. 2013; 38:111–123. [Бесплатная статья PMC] [PubMed] [Google Scholar]
117. Szyf M, Weaver IC, Champagne FA, Diorio J, Meaney MJ. Материнское программирование экспрессии и фенотипа стероидных рецепторов посредством метилирования ДНК у крыс. Передний нейроэндокринол. 2005; 26: 139–162. [PubMed] [Академия Google]
118. Шампанское Ф.А., Керли Дж.П. Эпигенетические механизмы, опосредующие долгосрочное влияние материнской заботы на развитие. Neurosci Biobehav Rev. 2009; 33: 593–600. [PubMed] [Google Scholar]
119. Фрейд С. Три эссе по теории сексуальности. В: Strachey TJ, редактор. Стандартное издание полного собрания психологических сочинений Зигмунда Фрейда.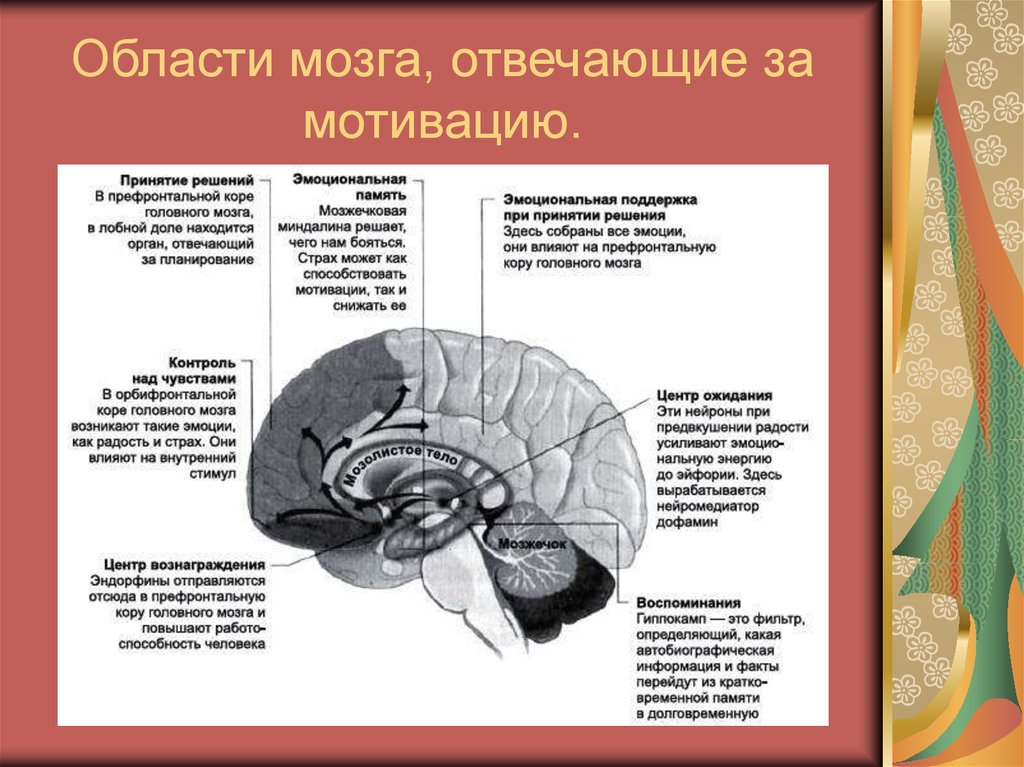 Том. 7. Хогарт Пресс; 1953. [Google Scholar]
Том. 7. Хогарт Пресс; 1953. [Google Scholar]
120. Кэмпбелл Б.А., Спир Н.Е. Онтогенез памяти. Psychol Rev. 1972; 79: 215–&. [PubMed] [Академия Google]
121. Копье Н.Е., Парсонс П. Анализ реактивационного лечения: Онтогенные детерминанты облегченного забывания. Лоуренс Эрлбаум Ассошиэйтс; Hillsdale, New Jersey: 1976. [Google Scholar]
122. Hayne H. Развитие детской памяти: последствия детской амнезии. Dev Rev. 2004; 24:33–73. [Google Scholar]
123. Терр Л. Что происходит с ранними воспоминаниями о травме? Исследование двадцати детей в возрасте до пяти лет во время задокументированных травмирующих событий. Журнал Американской академии детской и подростковой психиатрии. 1988;27:96–104. [PubMed] [Google Scholar]
124. Gaensbauer TJ. Травма в доречевой период. Симптомы, воспоминания и влияние на развитие. Психоаналитическое исследование ребенка. 1995; 50: 122–149. [PubMed] [Google Scholar]
125. Gaensbauer TJ, Jordan L. Психоаналитический взгляд на раннюю травму: интервью с тридцатью аналитиками, которые лечили взрослую жертву ограниченной травмы в раннем детстве. Журнал Американской психоаналитической ассоциации. 2009; 57: 947–977. [PubMed] [Академия Google]
Журнал Американской психоаналитической ассоциации. 2009; 57: 947–977. [PubMed] [Академия Google]
126. Peterson C, Rideout R. Воспоминание о неотложных состояниях, с которыми сталкиваются дети в возрасте 1 и 2 лет. Дев Психология. 1998; 34: 1059–1072. [PubMed] [Google Scholar]
127. Пейли Дж., Альперт Дж. Память о детской травме. Психоаналитическая психология. 2003; 20: 329–347. [Google Scholar]
128. Шеринга М., Генсбауэр Т. Посттравматическое стрессовое расстройство. В: Zeaanah C, редактор. Справочник по психическому здоровью младенцев. 2-е изд. Гилфорд Пресс; Нью-Йорк: 1999. С. 369–381. [Академия Google]
129. Терр ЛК. «Дикий ребенок»: Как три принципа исцеления организовали 12 лет психотерапии. Журнал Американской академии детской и подростковой психиатрии. 2003;42:1401–1409. [PubMed] [Google Scholar]
130. Terr LC. Травмы детства: план и обзор. Am J Психиатрия. 1991; 148:10–20. [PubMed] [Google Scholar]
Нейробиологи определили мозговую цепь, необходимую для формирования памяти | Новости Массачусетского технологического института
Когда мы навещаем друга или идем на пляж, наш мозг сохраняет кратковременную память об опыте в части мозга, называемой гиппокампом. Эти воспоминания позже «консолидируются», то есть переносятся в другую часть мозга для более длительного хранения.
Эти воспоминания позже «консолидируются», то есть переносятся в другую часть мозга для более длительного хранения.
Новое исследование нейронных цепей, лежащих в основе этого процесса, проведенное Массачусетским технологическим институтом, впервые показывает, что воспоминания на самом деле формируются одновременно в гиппокампе и в месте долговременного хранения в коре головного мозга. Однако долговременные воспоминания остаются «молчаливыми» около двух недель, прежде чем достигают зрелого состояния.
«Этот и другие открытия в этой статье обеспечивают комплексный механизм цепи для консолидации памяти», — говорит Сусуму Тонегава, профессор биологии и неврологии Пикауэра, директор Центра генетики нейронных цепей RIKEN-MIT в Институте Пиковера. Обучение и память, а также старший автор исследования.
Результаты, опубликованные в журнале Science от 6 апреля, могут потребовать некоторого пересмотра доминирующих моделей того, как происходит консолидация памяти, говорят исследователи.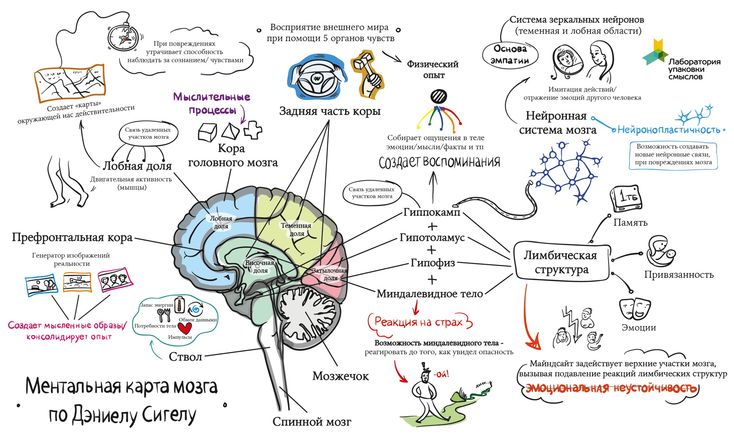
Ведущими авторами статьи являются ученый-исследователь Такаси Китамура, постдокторант Сачи Огава и аспирант Дирадж Рой. Другими авторами являются постдоки Терухиро Окуяма и Марк Моррисси, технический сотрудник Лилиан Смит и бывший постдок Роджер Редондо.
Долгосрочное хранение
Начавшиеся в 1950-х годах исследования знаменитого пациента с амнезией Генри Молисона, тогда известного как Пациент Х.М., показали, что гиппокамп необходим для формирования новых долговременных воспоминаний. Молисон, чей гиппокамп был поврежден во время операции, призванной помочь контролировать его эпилептические припадки, больше не мог хранить новые воспоминания после операции. Однако он все еще мог получить доступ к некоторым воспоминаниям, которые были сформированы до операции.
Это предполагает, что долгосрочные эпизодические воспоминания (воспоминания о конкретных событиях) хранятся вне гиппокампа. Ученые считают, что эти воспоминания хранятся в неокортексе, части мозга, которая также отвечает за когнитивные функции, такие как внимание и планирование.
Нейробиологи разработали две основные модели, описывающие перенос воспоминаний из кратковременной памяти в долговременную. Самая ранняя из них, известная как стандартная модель, предполагает, что краткосрочные воспоминания изначально формируются и хранятся только в гиппокампе, а затем постепенно передаются в долговременное хранилище в неокортексе и исчезают из гиппокампа.
Более поздняя модель, модель множественных следов, предполагает, что следы эпизодических воспоминаний остаются в гиппокампе. Эти следы могут хранить детали памяти, в то время как более общие контуры хранятся в неокортексе.
До недавнего времени не было возможности проверить эти теории. Большинство предыдущих исследований памяти были основаны на анализе того, как повреждение определенных областей мозга влияет на воспоминания. Однако в 2012 году лаборатория Тонегавы разработала способ маркировки клеток, называемых клетками энграмм, которые содержат определенные воспоминания. Это позволяет исследователям отслеживать схемы, участвующие в хранении и извлечении памяти. Они также могут искусственно реактивировать воспоминания с помощью оптогенетики — метода, который позволяет им включать или выключать клетки-мишени с помощью света.
Они также могут искусственно реактивировать воспоминания с помощью оптогенетики — метода, который позволяет им включать или выключать клетки-мишени с помощью света.
В новом исследовании Science исследователи использовали этот подход для маркировки клеток памяти у мышей во время события, вызывающего страх, то есть мягкого электрического удара, когда мышь находится в определенной камере. Затем они могли использовать свет для искусственной реактивации этих клеток памяти в разное время и посмотреть, вызывает ли эта реактивация поведенческую реакцию у мышей (замирание на месте). Исследователи также смогли определить, какие клетки памяти были активны, когда мышей поместили в камеру, где происходило обусловливание страха, побуждая их естественным образом вспоминать воспоминания.
Исследователи пометили клетки памяти в трех частях мозга: гиппокампе, префронтальной коре и базолатеральной миндалине, которая хранит эмоциональные ассоциации воспоминаний.
Всего через день после события, вызывающего страх, исследователи обнаружили, что воспоминания об этом событии хранятся в клетках инграмм как в гиппокампе, так и в префронтальной коре. Однако инграммные клетки в префронтальной коре были «молчаливыми» — они могли стимулировать замирание при искусственной активации светом, но не срабатывали во время естественного воспоминаний.
Однако инграммные клетки в префронтальной коре были «молчаливыми» — они могли стимулировать замирание при искусственной активации светом, но не срабатывали во время естественного воспоминаний.
«Префронтальная кора уже содержала определенную информацию о памяти, — говорит Китамура. «Это противоречит стандартной теории консолидации памяти, согласно которой воспоминания передаются постепенно. Память уже есть».
В течение следующих двух недель молчащие клетки памяти в префронтальной коре постепенно созревали, что отражалось в изменениях их анатомии и физиологической активности, пока клетки не стали необходимы животным для естественного воспоминания события. К концу того же периода инграммные клетки гиппокампа замолчали и больше не были нужны для естественного воспроизведения. Однако следы памяти остались: реактивация этих клеток светом все еще вызывала у животных замирание.
В базолатеральной миндалевидном теле после формирования воспоминаний клетки энграммы оставались неизменными на протяжении всего эксперимента.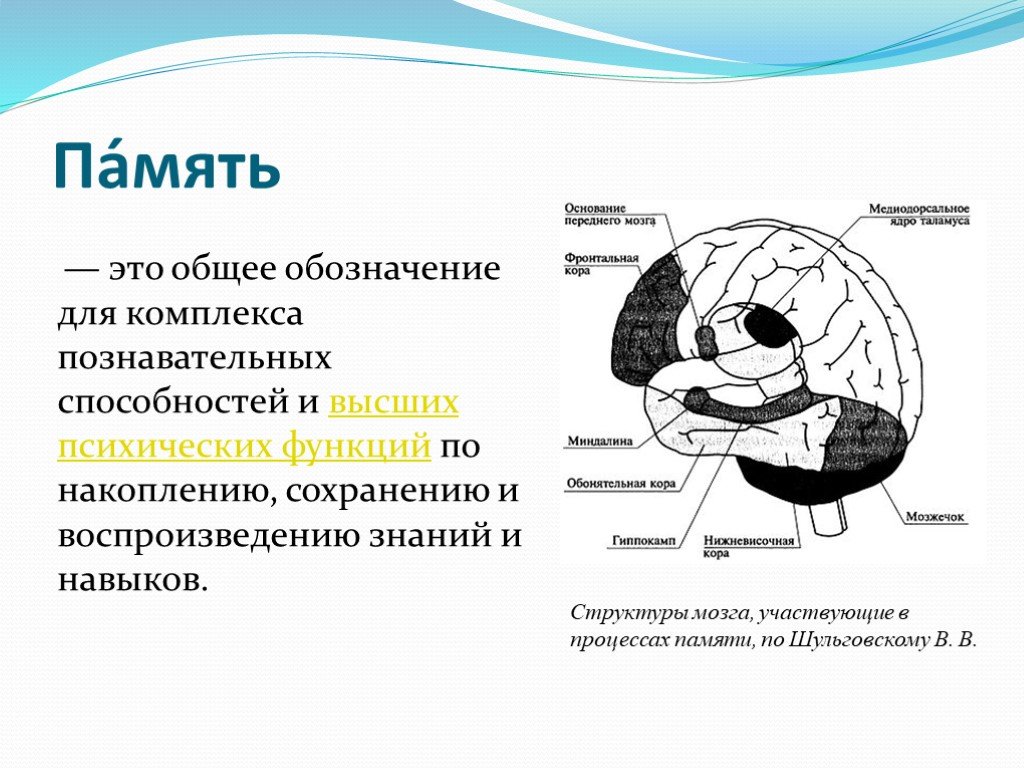 Те клетки, которые необходимы для того, чтобы вызвать эмоции, связанные с определенными воспоминаниями, взаимодействуют с инграммными клетками как в гиппокампе, так и в префронтальной коре.
Те клетки, которые необходимы для того, чтобы вызвать эмоции, связанные с определенными воспоминаниями, взаимодействуют с инграммными клетками как в гиппокампе, так и в префронтальной коре.
Пересмотр теории
Полученные данные свидетельствуют о том, что традиционные теории консолидации могут быть неточными, поскольку воспоминания формируются быстро и одновременно в префронтальной коре и гиппокампе в день тренировки.
«Они формируются параллельно, но потом идут разными путями. Префронтальная кора становится сильнее, а гиппокамп — слабее», — говорит Моррисси.
«Эта статья ясно показывает, что с самого начала инграммы формируются в префронтальной коре», — говорит Пол Франкленд, главный исследователь лаборатории нейробиологии в больнице для больных детей в Торонто, не участвовавший в исследовании. . «Это ставит под сомнение представление о том, что след памяти перемещается из гиппокампа в кору, и указывает на то, что эти цепи задействуются вместе в одно и то же время.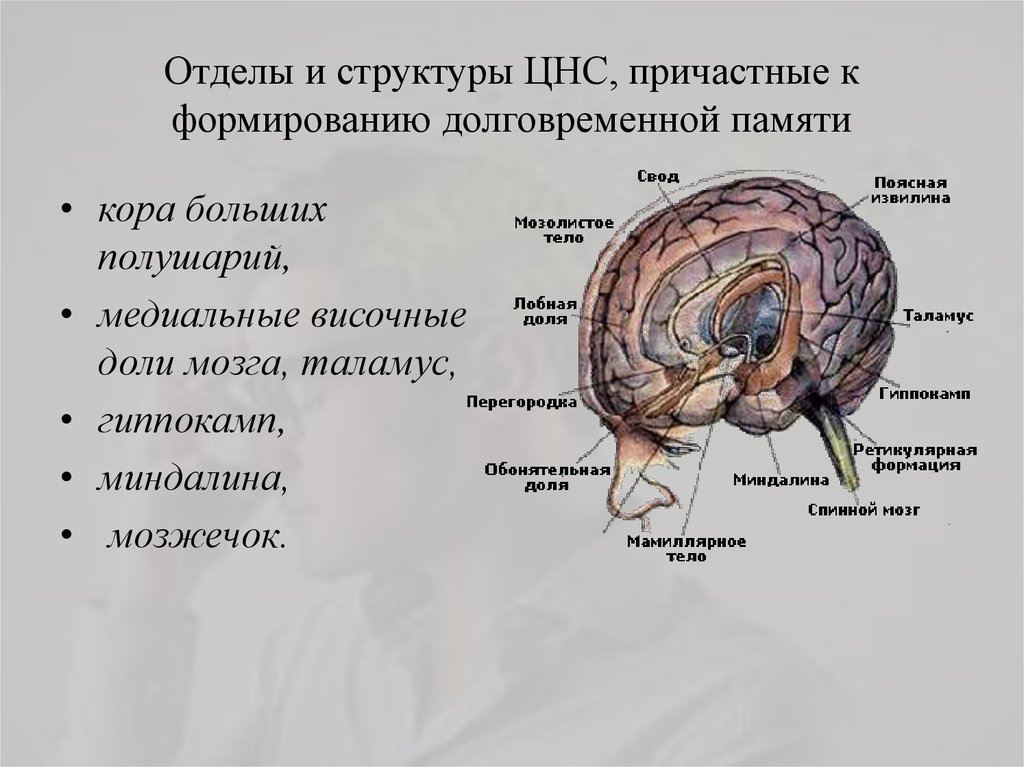 По мере старения воспоминаний происходит сдвиг в балансе того, какая цепь задействована при воспроизведении воспоминаний».
По мере старения воспоминаний происходит сдвиг в балансе того, какая цепь задействована при воспроизведении воспоминаний».
Необходимы дальнейшие исследования, чтобы определить, исчезают ли воспоминания из клеток гиппокампа полностью или остаются какие-то следы. Прямо сейчас исследователи могут наблюдать за клетками энграмм только около двух недель, но они работают над адаптацией своей технологии для работы в течение более длительного периода.
Китамура говорит, что, по его мнению, некоторые следы памяти могут оставаться в гиппокампе на неопределенный срок, сохраняя детали, которые извлекаются лишь изредка. «Чтобы различить два похожих эпизода, эта безмолвная инграмма может повторно активироваться, и люди могут восстановить подробную эпизодическую память даже в очень отдаленные моменты времени», — говорит он.
Исследователи также планируют продолжить изучение того, как происходит процесс созревания инграмм префронтальной коры. Это исследование уже показало, что связь между префронтальной корой и гиппокампом имеет решающее значение, потому что блокирование цепи, соединяющей эти две области, препятствует правильному созреванию корковых клеток памяти.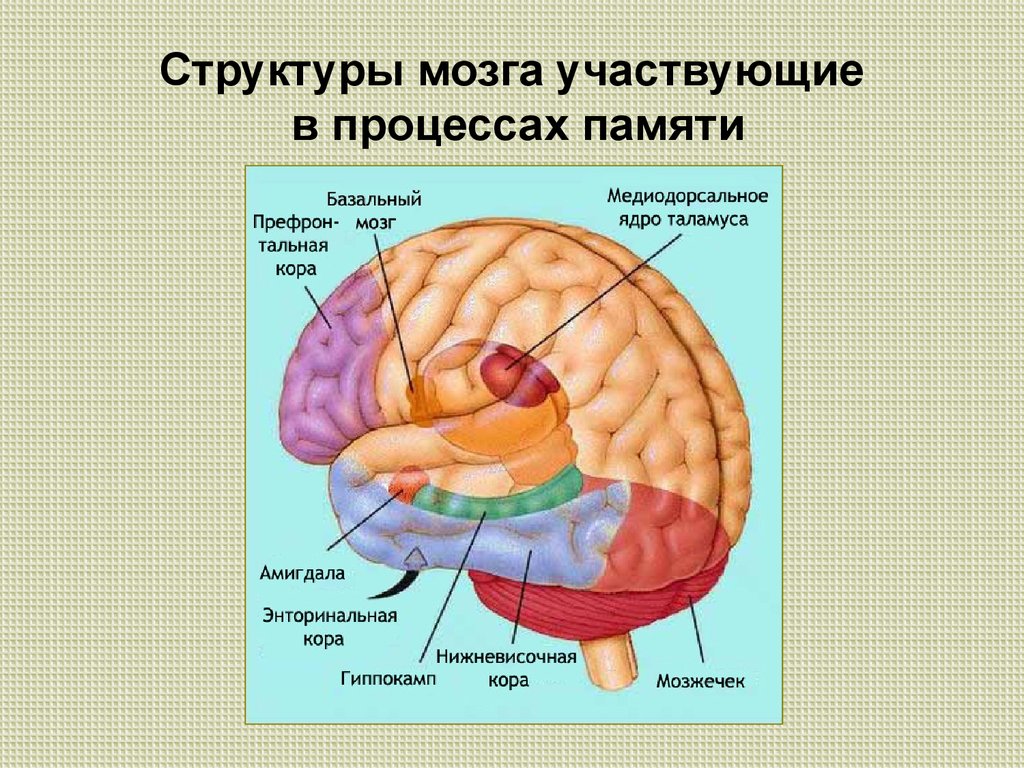
Исследование финансировалось Институтом изучения мозга RIKEN, Медицинским институтом Говарда Хьюза и Фондом JPB.
Поделиться этой новостной статьей:
Упоминания в прессе
Scientific American
Саймон Макин из Scientific American пишет, что исследователи Массачусетского технологического института обнаружили, что мозг использует дополнительную систему памяти, которая одновременно создает и хранит как долгосрочные, так и краткосрочные воспоминания. «Есть разделение труда. Гиппокамп может очень быстро формировать активные воспоминания, а кора головного мозга заботится о долговременной стабильности», — объясняет профессор Сусуму Тонегава.
Полная история через Scientific American →
PBS NOVA
Исследование, проведенное исследователем из Массачусетского технологического института, предполагает, что мозг одновременно создает долговременные и краткосрочные воспоминания, сообщает Тим Де Чант для NOVA .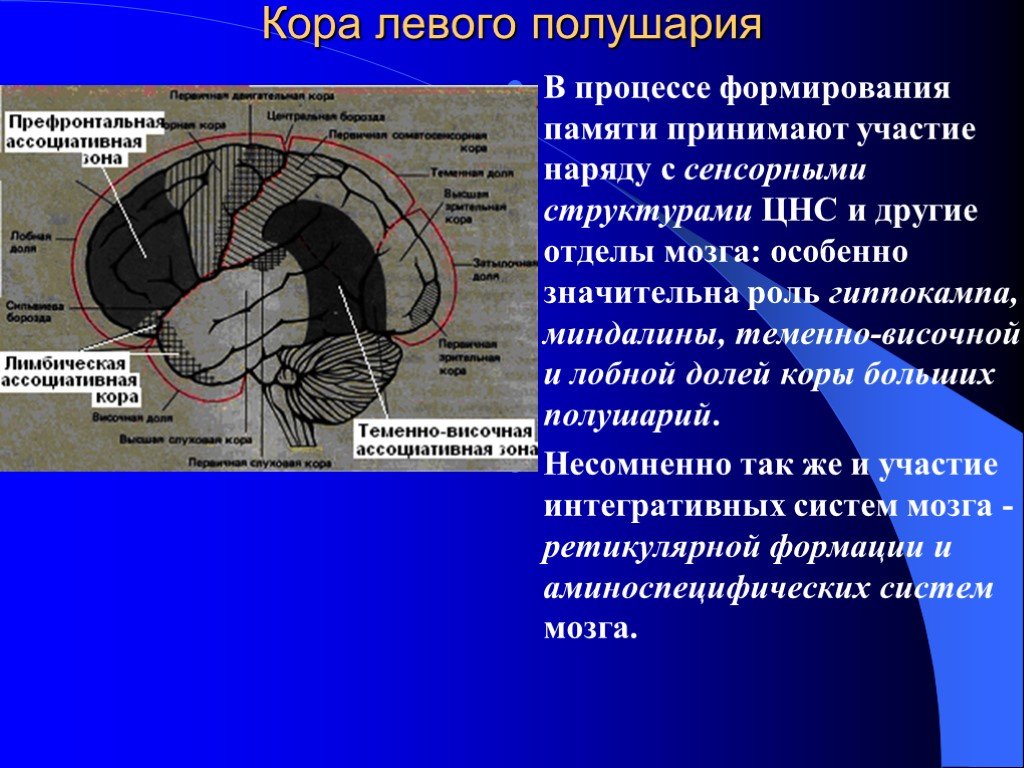 Полученные данные свидетельствуют о том, что одна версия «сохранена в гиппокампе, центре кратковременных воспоминаний, а другая хранится в коре головного мозга, где находятся наши долговременные воспоминания».
Полученные данные свидетельствуют о том, что одна версия «сохранена в гиппокампе, центре кратковременных воспоминаний, а другая хранится в коре головного мозга, где находятся наши долговременные воспоминания».
Полная история через PBS NOVA →
Boston Globe
Новое исследование, проведенное учеными Массачусетского технологического института, предполагает, что, вопреки предыдущим выводам, человеческий мозг может хранить краткосрочные и долгосрочные воспоминания одновременно, сообщает Энди Розен для Бостон глобус . Розен пишет, что результаты могут «привести к лучшему пониманию болезней памяти, таких как болезнь Альцгеймера».
Полная история через Boston Globe →
BBC News
Репортер BBC News Джеймс Галлахер пишет, что исследователи Массачусетского технологического института обнаружили, что мозг может одновременно создавать краткосрочные и долгосрочные версии воспоминаний.