Морфологический разбор слова «особенно»
Слово можно разобрать в 2 вариантах, в зависимости от того, в каком контексте оно используется.
1 вариант разбора
Часть речи: Наречие
2 вариант разбора
Часть речи: Краткое прилагательное
ОСОБЕННО — слово может быть как одушевленное так и неодушевленное, смотрите по предложению в котором оно используется.
Начальная форма слова: «ОСОБЕННЫЙ»
| Слово | Морфологические признаки |
|---|---|
| ОСОБЕННО |
|
Все формы слова ОСОБЕННО
ОСОБЕННЫЙ, ОСОБЕННОГО, ОСОБЕННОМУ, ОСОБЕННЫМ, ОСОБЕННОМ, ОСОБЕННАЯ, ОСОБЕННОЙ, ОСОБЕННУЮ, ОСОБЕННОЮ, ОСОБЕННОЕ, ОСОБЕННЫЕ, ОСОБЕННЫХ, ОСОБЕННЫМИ, ОСОБЕН, ОСОБЕННА, ОСОБЕННО, ОСОБЕННЫ, ОСОБЕННЕЕ, ОСОБЕННЕЙ, ПООСОБЕННЕЕ, ПООСОБЕННЕЙ
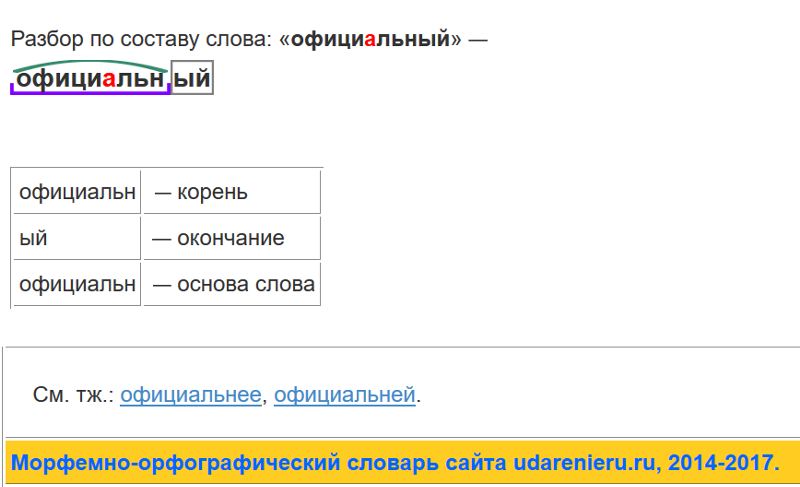
Разбор слова по составу особенно
особенно
| Основа слова | особенно |
|---|---|
| Корень | особ |
| Суффикс | енн |
| Суффикс | о |
Разобрать другие слова
Разбор слова в тексте или предложении
Если вы хотите разобрать слово «ОСОБЕННО» в конкретном предложении или тексте, то лучше использовать
морфологический разбор текста.
Найти синонимы к слову «особенно»
Примеры предложений со словом «особенно»
1
Вы удивлены, так как он не особенно умен, не особенно богат и не особенно интересен;
От полудня до полуночи, Эрих Мария Ремарк
2
Особенно талантливые, то есть особенно нервные, из оканчивающих школу остаются в ней в качестве воспитателей.
Ариэль, Александр Беляев, 1941г.
3
Адель дошла до того рокового предела, когда пустота личной жизни осознается особенно остро, становится особенно невыносимой.
На обломках трона, Евгений Маурин, 1899г.
4
Да, была причина, почему он мог быть или особенно счастлив, или особенно несчастлив.
Анна Каренина, Лев Толстой, 1878г.
5
Впоследствии узнал я, что именно в этом месте своей повести она особенно многое утаила и особенно многое передала несогласно с действительностью.
Огненный ангел, Валерий Брюсов, 1908-1909г.
Найти еще примеры предложений со словом ОСОБЕННО
Слова «деревянеть» морфологический и фонетический разбор
Объяснение правил деление (разбивки) слова «деревянеть» на слоги для переноса.
 ru поможет: фонетический и морфологический разобрать слово «деревянеть» по составу, правильно делить на слоги по провилам русского языка, выделить части слова, поставить ударение, укажет значение, синонимы, антонимы и сочетаемость к слову «деревянеть».
ru поможет: фонетический и морфологический разобрать слово «деревянеть» по составу, правильно делить на слоги по провилам русского языка, выделить части слова, поставить ударение, укажет значение, синонимы, антонимы и сочетаемость к слову «деревянеть».Содержимое:
- 1 Как перенести слово «деревянеть»
- 2 Предложения со словом «деревянеть»
- 3 Значение слова «деревянеть»
- 4 Как правильно пишется слово «деревянеть»
- 5 Спряжение глагола «деревянеть»
Как перенести слово «деревянеть»
де—ревянеть
дере—вянеть
деревя—неть
Предложения со словом «деревянеть»
На низменном месте в северной полосе от сырости воздуха и почвы побеги на зиму не успевают деревянеть и вымерзают зимою, на крепких же побегах ягоды поспевают поздно и бывают малосладкие; на юге низменное место, если на нём не бывает рос, может быть, напротив, благоприятным, особенно при засухе.
М. В. Рытов, Ягодники. Руководство по разведению крыжовника и смородины, 2012.
Значение слова «деревянеть»
ДЕРЕВЯНЕ’ТЬ. См. деревенеть. (Толковый словарь Ушакова)
Как правильно пишется слово «деревянеть»
Правописание слова «деревянеть»
Орфография слова «деревянеть»
Правильно слово пишется:
Нумерация букв в слове
Номера букв в слове «деревянеть» в прямом и обратном порядке:
Спряжение глагола «деревянеть»
Инфинитив
- деревянеть
Прошедшее время
- деревяне́л
Сослаг. наклонение
- деревяне́л бы
Изъявительное наклонение
я
деревяне́ю
ты
деревяне́ешь
он/она/оно
деревяне́ет
мы
деревяне́ем
вы
деревяне́ете
они
деревяне́ют
я
буду деревяне́ть
ты
будешь деревяне́ть
он/она/оно
будет деревяне́ть
мы
будем деревяне́ть
вы
будете деревяне́ть
они
будут деревянеть
я (он)
деревяне́л
я (она)
деревяне́ла
ты (он)
деревяне́л
ты (она)
деревяне́ла
он
деревяне́л
она
деревяне́ла
оно
деревяне́ло
мы
деревяне́ли
вы
деревяне́ли
они
деревяне́ли
Сослагательное наклонение
я (он)
деревяне́л бы
я (она)
деревяне́ла бы
ты (он)
деревяне́л бы
ты (она)
деревяне́ла бы
он
деревяне́л бы
она
деревяне́ла бы
оно
деревяне́ло бы
мы
деревяне́ли бы
вы
деревяне́ли бы
они
деревяне́ли бы
Повелительное наклонение
ты
деревяне́й
вы
деревяне́йте
Причастия
деревяне́ющий
деревяне́вший
деревяне́я
деревяне́в
Формирование, разборка и состав стрессовых гранул регулируются активностью АДФ-рибозилгидролазы альфавируса
. 2021 9 февраля; 118 (6): e2021719118.
2021 9 февраля; 118 (6): e2021719118.
doi: 10.1073/pnas.2021719118.
Аравинт Кумар Джаябалан 1 , Шриватсан Адиварахан 2 , Аакаш Коппула 3 , Рэйчи Абрахам 4 , Мона Батиш 3 5
, Даниэль Зенклузен 2 , Дайан Э. Гриффин 4 , Энтони К.Л. Леунг 6 7 8Принадлежности
- 1 Кафедра биохимии и молекулярной биологии, Школа общественного здравоохранения Блумберга, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205.

- 2 Департамент биохимии и молекулярной медицины, Университет Монреаля, Монреаль, QC h4T 1J4, Канада.
- 3 Факультет биологических наук, Делавэрский университет, Ньюарк, Делавэр, DE 19716.
- 4 W. Harry Feinstone Кафедра молекулярной микробиологии и иммунологии, Школа общественного здравоохранения Bloomberg, Университет Джона Хопкинса, Балтимор, MD 21205.
- 5 Кафедра медицинских и молекулярных наук, Делавэрский университет, Ньюарк, Делавэр, DE 19716.
- 6 Кафедра биохимии и молекулярной биологии, Школа общественного здравоохранения Блумберга, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205; anthony. [email protected].
- 7 Кафедра молекулярной биологии и генетики, Медицинский факультет Университета Джона Хопкинса, Балтимор, Мэриленд, 21205.
- 8 Кафедра онкологии, Медицинский факультет, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205.


- PMID: 33547245
- PMCID: PMC8017970
- DOI: 10.1073/пнас.2021719118
Бесплатная статья ЧВК
Аравинт Кумар Джаябалан и др.
Proc Natl Acad Sci U S A. .
.
Бесплатная статья ЧВК
. 2021 9 февраля; 118 (6): e2021719118.
doi: 10.1073/pnas.2021719118.
Авторы
Аравинт Кумар Джаябалан 1 , Шриватсан Адиварахан 2 , Аакаш Коппула 3 , Рэйчи Абрахам 4 , Мона Батиш 3 5 , Даниэль Зенклузен 2 , Дайан Э. Гриффин 4 , Энтони К.Л. Леунг 6 7 8
Принадлежности
- 1 Кафедра биохимии и молекулярной биологии, Школа общественного здравоохранения Блумберга, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205.
- 2 Департамент биохимии и молекулярной медицины, Университет Монреаля, Монреаль, QC h4T 1J4, Канада.
- 3 Факультет биологических наук, Делавэрский университет, Ньюарк, Делавэр, DE 19716.
- 4 W. Harry Feinstone Кафедра молекулярной микробиологии и иммунологии, Школа общественного здравоохранения Bloomberg, Университет Джона Хопкинса, Балтимор, MD 21205.
- 5 Кафедра медицинских и молекулярных наук, Делавэрский университет, Ньюарк, Делавэр, DE 19716.
- 6 Кафедра биохимии и молекулярной биологии, Школа общественного здравоохранения Блумберга, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205; anthony. [email protected].
- 7 Кафедра молекулярной биологии и генетики, Медицинский факультет Университета Джона Хопкинса, Балтимор, Мэриленд, 21205.
- 8 Кафедра онкологии, Медицинский факультет, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205.


- PMID: 33547245
- PMCID: PMC8017970
- DOI: 10.1073/пнас.2021719118
Абстрактный
Хотя биомолекулярные конденсаты стали важным биологическим явлением, механизмы, регулирующие их состав, и способы, которыми вирусы захватывают эти механизмы, остаются неясными. Альфавирусы, переносимые комарами, вызывают целый ряд заболеваний, от сыпи и артрита до энцефалита, при этом отсутствуют лицензированные лекарства для лечения или вакцины для профилактики. Неструктурный белок 3 фактора вирулентности альфавируса (nsP3) подавляет образование стрессовых гранул (SG) — класса цитоплазматических конденсатов, обогащенных факторами инициации трансляции и образующихся на ранней стадии инфекции. nsP3 имеет консервативный N-концевой макродомен, который гидролизует АДФ-рибозу из АДФ-рибозилированных белков, и С-концевой гипервариабельный домен, который связывает основной компонент SG G3BP1. Здесь мы показываем, что активность макродоменной гидролазы снижает АДФ-рибозилирование G3BP1, разбирает индуцированные вирусом SG и подавляет образование SG. Экспрессия nsP3 приводит к образованию отдельного класса конденсатов, в которых отсутствуют факторы инициации трансляции, но которые содержат G3BP1 и другие SG-ассоциированные РНК-связывающие белки. Экспрессия дефицитного по АДФ-рибозилгидролазе nsP3 приводит к образованию конденсатов, которые сохраняют факторы инициации трансляции, а также РНК-связывающие белки, сходные с SG.
Альфавирусы, переносимые комарами, вызывают целый ряд заболеваний, от сыпи и артрита до энцефалита, при этом отсутствуют лицензированные лекарства для лечения или вакцины для профилактики. Неструктурный белок 3 фактора вирулентности альфавируса (nsP3) подавляет образование стрессовых гранул (SG) — класса цитоплазматических конденсатов, обогащенных факторами инициации трансляции и образующихся на ранней стадии инфекции. nsP3 имеет консервативный N-концевой макродомен, который гидролизует АДФ-рибозу из АДФ-рибозилированных белков, и С-концевой гипервариабельный домен, который связывает основной компонент SG G3BP1. Здесь мы показываем, что активность макродоменной гидролазы снижает АДФ-рибозилирование G3BP1, разбирает индуцированные вирусом SG и подавляет образование SG. Экспрессия nsP3 приводит к образованию отдельного класса конденсатов, в которых отсутствуют факторы инициации трансляции, но которые содержат G3BP1 и другие SG-ассоциированные РНК-связывающие белки. Экспрессия дефицитного по АДФ-рибозилгидролазе nsP3 приводит к образованию конденсатов, которые сохраняют факторы инициации трансляции, а также РНК-связывающие белки, сходные с SG. Таким образом, наши данные показывают, что АДФ-рибозилирование контролирует состав биомолекулярных конденсатов, особенно локализацию факторов инициации трансляции, во время инфекции альфавирусом.
Таким образом, наши данные показывают, что АДФ-рибозилирование контролирует состав биомолекулярных конденсатов, особенно локализацию факторов инициации трансляции, во время инфекции альфавирусом.
Ключевые слова: АДФ-рибозилирование; альфавирус; биомолекулярные конденсаты; макродомен; стрессовые гранулы.
Заявление о конфликте интересов
Авторы заявляют об отсутствии конкурирующих интересов.
Цифры
Рис. 1.
Два различных класса биомолекулярных…
Рис. 1.
При инфицировании альфавирусом образуются два различных класса биомолекулярных конденсатов. ( А )…
( А )…
При инфицировании альфавирусом образуются два различных класса биомолекулярных конденсатов. ( A ) Клетки U2OS, стабильно экспрессирующие GFP-eIF3g, были инфицированы WT CHIKV mCherry при MOI 10, подвергались визуализации живых клеток с интервалом времени 12 минут в течение 14 часов. Показанные изображения представляют собой снимки различных стадий заражения. ( B ) Клетки U2OS были либо ложно инфицированы, либо инфицированы CHIKV. Через двенадцать часов после инфицирования клетки лизировали и блоттировали против указанных антител. ( C ) Скрипичный график показывает распределение временных точек (минут), представляющих появление, исчезновение SG и появление конденсата nsP3. Были включены 97 клеток из трех независимых экспериментов, и в каждой клетке были измерены моменты времени появления/исчезновения СГ и появления конденсатов nsP3. Столбики погрешностей соответствуют SD. ( D ) Клетки U2OS инфицировали CHIKV WT при множественности заражения 1. Через 6 ч после заражения (hpi) клетки фиксировали и иммуноокрашивали на nsP3 и определяли белки SG. ( E ) Клетки U2OS, трансфицированные GFP-меченым nsP3 WT , либо не подвергали стрессу, либо обрабатывали 100 мкг/мл циклогексимида (CHX), 0,2 мМ арсенита отдельно (ARS), либо совместно обрабатывали 0,2 мМ арсенита и 100 мкг/мл циклогексимид (ARS+CHX) в течение 30 мин. Затем клетки фиксировали и иммуноокрашивали на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( F ) Гистограмма показывает процент GFP-положительных клеток с SG из трех независимых экспериментов. Столбики погрешностей соответствуют среднему значению ± стандартное отклонение. (Шкала, 10 мкм.)
( D ) Клетки U2OS инфицировали CHIKV WT при множественности заражения 1. Через 6 ч после заражения (hpi) клетки фиксировали и иммуноокрашивали на nsP3 и определяли белки SG. ( E ) Клетки U2OS, трансфицированные GFP-меченым nsP3 WT , либо не подвергали стрессу, либо обрабатывали 100 мкг/мл циклогексимида (CHX), 0,2 мМ арсенита отдельно (ARS), либо совместно обрабатывали 0,2 мМ арсенита и 100 мкг/мл циклогексимид (ARS+CHX) в течение 30 мин. Затем клетки фиксировали и иммуноокрашивали на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( F ) Гистограмма показывает процент GFP-положительных клеток с SG из трех независимых экспериментов. Столбики погрешностей соответствуют среднему значению ± стандартное отклонение. (Шкала, 10 мкм.)
Рис. 2.
Макродомен nsP3 подавляет образование СГ. …
…
Рис. 2.
Макродомен nsP3 подавляет образование СГ. ( A ) Схематическое изображение nsP3…
Рис. 2. Макродомен nsP3 подавляет образование СГ. ( A ) Схематическое представление архитектуры домена nsP3. Клетки U2OS, трансфицированные GFP-мечеными nsP3 MD , nsP3 ZBD или nsP3 HVD , либо обрабатывали 0,2 мМ арсенита отдельно, либо совместно обрабатывали 100 мкг/мл циклогексимида в течение 30 мин. Затем клетки обрабатывали и иммуноокрашивали на маркеры SG G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( B ) Гистограмма показывает процент GFP-позитивных клеток с SG. *** P < 0,005, двусторонний, непарный тест Стьюдента t . Столбики погрешностей соответствуют среднему значению ± стандартное отклонение для трех независимых экспериментов. ( C ) Клетки U2OS, трансфицированные вектором GFP, nsP3 WT с меткой GFP или nsP3 AGDA , обрабатывали 0,2 мМ арсенита в течение 30 минут и подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( D ) Гистограмма показывает процент GFP-позитивных клеток с SG. * P < 0,05, *** P < 0,001, двусторонний, непарный тест Стьюдента t . Планки погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. (Шкала шкалы, 10 мкм.)
( C ) Клетки U2OS, трансфицированные вектором GFP, nsP3 WT с меткой GFP или nsP3 AGDA , обрабатывали 0,2 мМ арсенита в течение 30 минут и подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( D ) Гистограмма показывает процент GFP-позитивных клеток с SG. * P < 0,05, *** P < 0,001, двусторонний, непарный тест Стьюдента t . Планки погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. (Шкала шкалы, 10 мкм.)
Рис. 3.
АДФ-рибозилгидролазная активность nsP3 подавляет…
Рис. 3.
АДФ-рибозилгидролазная активность nsP3 подавляет образование СГ. ( A ) Трансфецированные клетки U2OS…
Рис. 3.
3. АДФ-рибозилгидролазная активность nsP3 подавляет образование SG. ( A ) Клетки U2OS, трансфицированные GFP-меченым nsP3 WT или другими точечными мутантами nsP3 (D10A, G32E, G32S, G112E и Y114A), обрабатывали 0,2 мМ арсенита в течение 30 минут и подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( B ) Клетки 293F трансфицировали либо вектором GFP, либо меченным GFP nsP3 WT , nsP3 G32E , nsP3 G32S или nsP3 Y114A . После 24-часовой трансфекции клетки осаждали, лизировали, иммунопреципитировали с использованием антител к GFP и проводили блоттинг иммунопреципитатов против антител G3BP1 и GFP. ( C ) Гистограмма показывает процент GFP-положительных клеток с SG в A . ( Нижний ) Связывание АДФ-рибозы и гидролазная активность протестированных мутантов (31). ** P < 0,01, двусторонний, непарный Стьюдента т тест. Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( D ) Клетки, трансфицированные GFP-мечеными nsP3 WT или nsP3 G32E , совместно обрабатывали 0,2 мМ арсенита и 100 мкг/мл циклогексимида для получения 30-минимина. Затем клетки подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( E ) В таблице обобщена колокализация nsP3 (WT или G32E) с различными связывающими мРНК белками и факторами трансляции, как показано на Приложение SI , рис. S3 A . ( F ) Микроскопия со сверхвысоким разрешением показывает, как распределение сигнала поли(А)+ мРНК перекрывается с мутантным nsP3 WT или G32E. Клетки U2OS, трансфицированные меченым GFP nsP3 WT или nsP3 G32E , обрабатывали 0,2 мМ арсенита в течение 30 минут и гибридизовали с олиго(dT) зондами с последующим окрашиванием на GFP. Зеленые и красные прямоугольники обозначают конденсаты из nsP3-трансфицированных и нетрансфицированных клеток соответственно.
Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( D ) Клетки, трансфицированные GFP-мечеными nsP3 WT или nsP3 G32E , совместно обрабатывали 0,2 мМ арсенита и 100 мкг/мл циклогексимида для получения 30-минимина. Затем клетки подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( E ) В таблице обобщена колокализация nsP3 (WT или G32E) с различными связывающими мРНК белками и факторами трансляции, как показано на Приложение SI , рис. S3 A . ( F ) Микроскопия со сверхвысоким разрешением показывает, как распределение сигнала поли(А)+ мРНК перекрывается с мутантным nsP3 WT или G32E. Клетки U2OS, трансфицированные меченым GFP nsP3 WT или nsP3 G32E , обрабатывали 0,2 мМ арсенита в течение 30 минут и гибридизовали с олиго(dT) зондами с последующим окрашиванием на GFP. Зеленые и красные прямоугольники обозначают конденсаты из nsP3-трансфицированных и нетрансфицированных клеток соответственно. (Шкала баров, 10 мкм.)
(Шкала баров, 10 мкм.)
Рис. 4.
Дифференциальная способность связывания PAR и АДФ-рибозилирование…
Рис. 4.
Дифференциальная ФАР-связывающая способность и АДФ-рибозилирование компонентов СГ. ( A ) 293F ячейки…
Рис. 4. Дифференциальная ФАР-связывающая способность и АДФ-рибозилирование компонентов СГ. ( 9Клетки 0179 A ) 293F трансфицировали либо вектором GFP, либо меченными GFP G3BP1, G3BP2, eIF3d, eIF3g или eIF3i в течение 36 часов. Затем клетки осаждали, лизировали и инкубировали со 100 пмоль биотина-PAR и гранулами стрептавидина. Затем пулдаун стрептавидина блоттировали антителами против GFP. ( B ) Клетки 293F, трансфицированные либо вектором GFP, либо мечеными GFP G3BP1, G3BP2, eIF3d, eIF3g или eIF3i в течение 36 часов. Затем клетки осаждали, лизировали и подвергали иммунопреципитации с использованием антител против GFP. Затем иммунопреципитаты блоттировали реагентом pan-ADPr.
Затем клетки осаждали, лизировали и подвергали иммунопреципитации с использованием антител против GFP. Затем иммунопреципитаты блоттировали реагентом pan-ADPr.
Рис. 5.
nsP3 снижает АДФ-рибозилирование…
Рис. 5.
nsP3 снижает АДФ-рибозилирование основного компонента SG G3BP1. ( А ) 293F…
Рис. 5. nsP3 снижает АДФ-рибозилирование основного компонента SG G3BP1. ( 9Клетки 0179 A ) 293F трансфицировали либо GFP, G3BP1, меченым GFP, либо котрансфицировали G3BP1, меченым GFP, с возрастающей концентрацией меченого FLAG nsP3 WT . После 24-часовой трансфекции клетки осаждали, лизировали, иммунопреципитировали с использованием антител против GFP и иммуноблоттировали с реагентом pan-ADPr. ( B ) Клетки 293F трансфицировали GFP, G3BP1, меченным GFP отдельно, или котрансфицировали G3BP1, меченым GFP, либо с меченым FLAG nsP3 WT , либо с nsP3 G32E . Через 24 часа после трансфекции клетки осаждали, лизировали, подвергали иммунопреципитации с использованием антител против GFP и иммуноблотингу с реагентом pan-ADPr. ( C ) Клетки 293F, трансфицированные меченным GFP G3BP1, лизировали и подвергали иммунопреципитации с использованием антител против GFP. Иммунопреципитаты разделяли на две половины и инкубировали либо с буфером, либо с 5 мкг ЧИКВ МД в течение 1 ч при 37 °С. После инкубации шарики промывали и промокали реагентом pan-ADPr. ( D ) Схематическое изображение доменной структуры G3BP1. ( E – G ) Клетки с двойным нокаутом (dKO) U2OS G3BP1/2 трансфицировали либо вектором GFP, либо конструкциями G3BP1, меченными GFP, в течение 36 часов. ( E ) Затем клетки либо лизировали, иммунопреципитировали с использованием антител против GFP и подвергали блотингу с реагентом pan-ADPr (звездочки обозначают тяжелую и легкую цепи), либо ( F ) пермеабилизировали, фиксировали и иммуноокрашивали для PABP (красный) и eIF3b (синий).
( B ) Клетки 293F трансфицировали GFP, G3BP1, меченным GFP отдельно, или котрансфицировали G3BP1, меченым GFP, либо с меченым FLAG nsP3 WT , либо с nsP3 G32E . Через 24 часа после трансфекции клетки осаждали, лизировали, подвергали иммунопреципитации с использованием антител против GFP и иммуноблотингу с реагентом pan-ADPr. ( C ) Клетки 293F, трансфицированные меченным GFP G3BP1, лизировали и подвергали иммунопреципитации с использованием антител против GFP. Иммунопреципитаты разделяли на две половины и инкубировали либо с буфером, либо с 5 мкг ЧИКВ МД в течение 1 ч при 37 °С. После инкубации шарики промывали и промокали реагентом pan-ADPr. ( D ) Схематическое изображение доменной структуры G3BP1. ( E – G ) Клетки с двойным нокаутом (dKO) U2OS G3BP1/2 трансфицировали либо вектором GFP, либо конструкциями G3BP1, меченными GFP, в течение 36 часов. ( E ) Затем клетки либо лизировали, иммунопреципитировали с использованием антител против GFP и подвергали блотингу с реагентом pan-ADPr (звездочки обозначают тяжелую и легкую цепи), либо ( F ) пермеабилизировали, фиксировали и иммуноокрашивали для PABP (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( G ) Гистограмма показывает процент GFP-позитивных клеток с SG. нс, незначащий, двусторонний, непарный Стьюдента т тест. Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( H и I ) Клетки U2OS G3BP1/2 dKO котрансфицировали в двух комбинациях: 1) FLAG-вектор с G3BP1 или G3BP1-меченым ∆1– 168, и 2) nsP3 с маркировкой FLAG с G3BP1 или G3BP1 с меткой GFP ∆1–168. Через 36 ч после трансфекции клетки фиксировали и окрашивали на FLAG (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. Гистограмма показывает процент GFP-положительных клеток с SG. *** Р < 0,001, **** P < 0,0001, двусторонний, непарный Критерий Стьюдента t . Планки погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. (Шкала шкалы, 10 мкм.)
Звездочки указывают на нетрансфицированные клетки. ( G ) Гистограмма показывает процент GFP-позитивных клеток с SG. нс, незначащий, двусторонний, непарный Стьюдента т тест. Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( H и I ) Клетки U2OS G3BP1/2 dKO котрансфицировали в двух комбинациях: 1) FLAG-вектор с G3BP1 или G3BP1-меченым ∆1– 168, и 2) nsP3 с маркировкой FLAG с G3BP1 или G3BP1 с меткой GFP ∆1–168. Через 36 ч после трансфекции клетки фиксировали и окрашивали на FLAG (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. Гистограмма показывает процент GFP-положительных клеток с SG. *** Р < 0,001, **** P < 0,0001, двусторонний, непарный Критерий Стьюдента t . Планки погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. (Шкала шкалы, 10 мкм.)
Рис. 6.
6.
АДФ-рибозилгидролазная активность nsP3 подавляет…
Рис. 6.
АДФ-рибозилгидролазная активность nsP3 подавляет образование СГ при альфавирусной инфекции. ( А )…
Рис. 6. АДФ-рибозилгидролазная активность nsP3 подавляет образование SG при альфавирусной инфекции. ( A ) Титры вируса WT и мутанта Y114A в клетках NSC34 (31) и U2OS проанализированы при 24 hpi. Данные представлены в виде единиц образования бляшек (БОЕ)/мл ± SEM. ( B ) Клетки 293T, трансфицированные вектором GFP или GFP-G3BP1 в течение 12 часов, были либо ложно инфицированы, либо инфицированы CHIKV WT или CHIKV Y114A при МВД 1 в течение 24 часов. При 24 HPI клетки лизировали, иммунопреципитировали с использованием антител против GFP и блоттировали реагентом pan-ADPr. ( C и D ) Клетки U2OS инфицировали либо CHIKV WT , либо CHIKV Y114A при множественности заражения 1. При 5,5 hpi клетки обрабатывали 0,2 мМ арсенита в течение 30 минут и окрашивали иммуноокрашиванием на nsP3 (зеленый) , G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. Гистограмма показывает процент инфицированных вирусом клеток (nsP3-позитивных) с SG. ** P < 0,01, двусторонний, непарный тест Стьюдента t . Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( E ) Клетки U2OS были инфицированы либо CHIKV WT , либо CHIKV Y114A при MOI 5. При 7,5 HPI клетки обрабатывали 0,2 мМ арсенита. в течение 30 мин. Затем обработанные клетки фиксировали и гибридизовали с зондами вРНК (красные) с последующим иммуноокрашиванием на G3BP1 (зеленый) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( Ф ) Прямоугольная диаграмма показывает процент клеток с вРНК , совместно локализованных с eIF3b/G3BP1 , на рис. 6 E и SI Приложение , рис.
При 5,5 hpi клетки обрабатывали 0,2 мМ арсенита в течение 30 минут и окрашивали иммуноокрашиванием на nsP3 (зеленый) , G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. Гистограмма показывает процент инфицированных вирусом клеток (nsP3-позитивных) с SG. ** P < 0,01, двусторонний, непарный тест Стьюдента t . Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( E ) Клетки U2OS были инфицированы либо CHIKV WT , либо CHIKV Y114A при MOI 5. При 7,5 HPI клетки обрабатывали 0,2 мМ арсенита. в течение 30 мин. Затем обработанные клетки фиксировали и гибридизовали с зондами вРНК (красные) с последующим иммуноокрашиванием на G3BP1 (зеленый) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( Ф ) Прямоугольная диаграмма показывает процент клеток с вРНК , совместно локализованных с eIF3b/G3BP1 , на рис. 6 E и SI Приложение , рис. S5 E . Около 20 клеток из каждой временной точки были количественно определены для колокализации. * P < 0,05, нс, недостоверный, двусторонний, непарный критерий Стьюдента t . (Шкала, 10 мкм.)
S5 E . Около 20 клеток из каждой временной точки были количественно определены для колокализации. * P < 0,05, нс, недостоверный, двусторонний, непарный критерий Стьюдента t . (Шкала, 10 мкм.)
Рис. 7.
АДФ-рибозилгидролазная активность nsP3 регулирует…
Рис. 7.
АДФ-рибозилгидролазная активность nsP3 регулирует разборку SG. ( A ) Схематическое изображение…
Рис. 7. АДФ-рибозилгидролазная активность nsP3 регулирует разборку SG. ( A ) Схематическое изображение установки эксперимента по визуализации живых клеток. ( B и C ) Клетки U2OS, стабильно экспрессирующие GFP-eIF3g, были инфицированы WT или Y114A CHIKV 9.0007 mCherry с MOI 10 и подвергали визуализации живых клеток с интервалом времени 12 минут в течение 14 часов. Показанные изображения представляют собой моменты времени, когда SG не наблюдались (t = -12 мин), за которыми следует момент времени, когда наблюдались SG (t = 0 ч). Белые стрелки указывают на СГ. (Масштабная линейка, 10 мкм.) Скрипичный график показывает распределение среднего времени пребывания SG (минуты) в клетках, инфицированных WT и Y114A CHIKV mCherry . *** P < 0,001, двусторонний, непарный Стьюдента т тест. WT ( n = 51) и Y114A ( n = 77) инфицированные клетки из трех независимых экспериментов были включены для измерения времени пребывания SG. Столбики погрешностей соответствуют SD. ( D ) Модель сборки и разборки SG, индуцированной вирусной инфекцией, регулируемой активностью nsP3 ADP-рибозилгидролазы.
Показанные изображения представляют собой моменты времени, когда SG не наблюдались (t = -12 мин), за которыми следует момент времени, когда наблюдались SG (t = 0 ч). Белые стрелки указывают на СГ. (Масштабная линейка, 10 мкм.) Скрипичный график показывает распределение среднего времени пребывания SG (минуты) в клетках, инфицированных WT и Y114A CHIKV mCherry . *** P < 0,001, двусторонний, непарный Стьюдента т тест. WT ( n = 51) и Y114A ( n = 77) инфицированные клетки из трех независимых экспериментов были включены для измерения времени пребывания SG. Столбики погрешностей соответствуют SD. ( D ) Модель сборки и разборки SG, индуцированной вирусной инфекцией, регулируемой активностью nsP3 ADP-рибозилгидролазы.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
Роль Chikungunya nsP3 в регуляции активности G3BP1, формировании стрессовых гранул и эффективности лекарств.
Лу Х, Алам У, Уиллис С, Кеннеди Д. Лу X и др. Арх Мед Рез. 2021 Январь; 52(1):48-57. doi: 10.1016/j.arcmed.2020.10.002. Epub 2020 31 октября. Арх Мед Рез. 2021. PMID: 33131924
Компоненты стрессовых гранул G3BP1 и G3BP2 играют провирусную роль на ранней стадии репликации вируса чикунгунья.
Шольте Ф.Е., Тас А., Албулеску И.С., Жусинайте Э., Меритс А., Снайдер Э.Дж., ван Хемерт М.Дж. Шольте Ф.Е. и соавт. Дж Вирол. 2015 г., апрель 89(8):4457-69. doi: 10.1128/ОВИ.03612-14. Epub 2015 4 февраля. Дж Вирол. 2015. PMID: 25653451 Бесплатная статья ЧВК.
Отдельные домены G3BP способствуют эффективной кластеризации комплексов репликации альфавируса и задействованию аппарата инициации трансляции.
Гётте Б.
, Панас М.Д., Хеллстрем К., Лю Л., Самрин Б., Ларссон О., Ахола Т., Макинерни Г.М.
Гётте Б. и др.
PLoS Патог. 14 июня 2019 г .; 15 (6): e1007842. doi: 10.1371/journal.ppat.1007842. Электронная коллекция 2019 июнь.
PLoS Патог. 2019.
PMID: 31199850
Бесплатная статья ЧВК.Прогресс исследований структуры и функции G3BP.
Кан В, Ван И, Ян В, Чжан Дж, Чжэн Х, Ли Д. Канг В. и др. Фронт Иммунол. 2021 30 авг;12:718548. дои: 10.3389/fimmu.2021.718548. Электронная коллекция 2021. Фронт Иммунол. 2021. PMID: 34526993 Бесплатная статья ЧВК. Обзор.
Распутин десять лет спустя, а распущенность больше, чем когда-либо? Обзор G3BP.
Алам Ю., Кеннеди Д. Алам У и др. Biochim Biophys Acta Mol Cell Res.
2019 март; 1866 (3): 360-370. doi: 10.1016/j.bbamcr.2018.09.001. Epub 2018 5 сентября.
Biochim Biophys Acta Mol Cell Res. 2019.
PMID: 30595162
Бесплатная статья ЧВК.
Обзор.

 , Панас М.Д., Хеллстрем К., Лю Л., Самрин Б., Ларссон О., Ахола Т., Макинерни Г.М.
Гётте Б. и др.
PLoS Патог. 14 июня 2019 г .; 15 (6): e1007842. doi: 10.1371/journal.ppat.1007842. Электронная коллекция 2019 июнь.
PLoS Патог. 2019.
PMID: 31199850
Бесплатная статья ЧВК.
, Панас М.Д., Хеллстрем К., Лю Л., Самрин Б., Ларссон О., Ахола Т., Макинерни Г.М.
Гётте Б. и др.
PLoS Патог. 14 июня 2019 г .; 15 (6): e1007842. doi: 10.1371/journal.ppat.1007842. Электронная коллекция 2019 июнь.
PLoS Патог. 2019.
PMID: 31199850
Бесплатная статья ЧВК. 2019 март; 1866 (3): 360-370. doi: 10.1016/j.bbamcr.2018.09.001. Epub 2018 5 сентября.
Biochim Biophys Acta Mol Cell Res. 2019.
PMID: 30595162
Бесплатная статья ЧВК.
Обзор.
2019 март; 1866 (3): 360-370. doi: 10.1016/j.bbamcr.2018.09.001. Epub 2018 5 сентября.
Biochim Biophys Acta Mol Cell Res. 2019.
PMID: 30595162
Бесплатная статья ЧВК.
Обзор.Посмотреть все похожие статьи
Цитируется
Отдельная фаза? Поли(АДФ-рибоза) против РНК в организации биомолекулярных конденсатов.
Алемасова Е.Е., Лаврик О.И. Алемасова Е.Е. и соавт. Нуклеиновые Кислоты Res. 2022 28 октября; 50 (19): 10817-10838. doi: 10.1093/nar/gkac866. Нуклеиновые Кислоты Res. 2022. PMID: 36243979 Бесплатная статья ЧВК. Обзор.
Ингибирование cGAS ослабляет индуцированные Alu РНК иммунные ответы и цитотоксичность в пигментированном эпителии сетчатки.
Li J, Zhang F, Bian W, Chen Y, Liu J, Liu Z, Xiong Y, Wan X.
Ли Дж. и др.
Клетка Биоски. 2022 25 июля; 12 (1): 116. doi: 10.1186/s13578-022-00854-y.
Клетка Биоски. 2022.
PMID: 35879806
Бесплатная статья ЧВК.Стрессовые шаги: прогресс и проблемы в понимании вызванной стрессом конденсации и накопления мРНК в стрессовых гранулах.
Glauninger H, Wong Hickernell CJ, Bard JAM, Drummond DA. Глаунингер Х. и др. Мол Ячейка. 2022 21 июля; 82 (14): 2544-2556. doi: 10.1016/j.molcel.2022.05.014. Epub 2022 3 июня. Мол Ячейка. 2022. PMID: 35662398 Обзор.
Роль стресс-гранул в подавлении репликации вируса эндорибонуклеазой вируса инфекционного бронхита.
Чжао Дж., Фэн Д., Чжао Й., Хуан М., Чжан С., Чжан Г. Чжао Дж. и др. Дж Вирол. 22 июня 2022 г .;96(12):e0068622.
doi: 10.1128/jvi.00686-22. Epub 2022 31 мая.
Дж Вирол. 2022.
PMID: 35638780Внутриклеточные моно-АДФ-рибозилтрансферазы в интерфазе вирус-хозяин.
Люшер Б., Верхейрстратен М., Криг С., Корн П. Люшер Б. и соавт. Cell Mol Life Sci. 2022 10 мая; 79(6): 288. doi: 10.1007/s00018-022-04290-6. Cell Mol Life Sci. 2022. PMID: 35536484 Бесплатная статья ЧВК. Обзор.
 Ли Дж. и др.
Клетка Биоски. 2022 25 июля; 12 (1): 116. doi: 10.1186/s13578-022-00854-y.
Клетка Биоски. 2022.
PMID: 35879806
Бесплатная статья ЧВК.
Ли Дж. и др.
Клетка Биоски. 2022 25 июля; 12 (1): 116. doi: 10.1186/s13578-022-00854-y.
Клетка Биоски. 2022.
PMID: 35879806
Бесплатная статья ЧВК. doi: 10.1128/jvi.00686-22. Epub 2022 31 мая.
Дж Вирол. 2022.
PMID: 35638780
doi: 10.1128/jvi.00686-22. Epub 2022 31 мая.
Дж Вирол. 2022.
PMID: 35638780Просмотреть все статьи «Цитируется по»
Типы публикаций
термины MeSH
вещества
Грантовая поддержка
- R01 GM104135/GM/NIGMS NIH HHS/США
- R56 AI137264/AI/NIAID NIH HHS/США
- PJT-148932/CIHR/Канада
- UDRF_SI_19A00244/CIHR/Канада
Состав против Наследства: Как выбрать?
В начале.
 ..
..… не было ни наследования, ни композиции, только код.
И код был громоздким, повторяющимся, блочным, несчастливым, многословным и усталым.
Копирование и вставка были основными механизмами повторного использования кода. Процедуры и функции были редкостью, к новомодным гаджетам относились с подозрением. Вызов процедуры был дорогим! Отделение кусков кода от основной логики вызвало путаницу!
Темное время было.
Затем мир осветил свет объектно-ориентированного программирования (ООП)… И мир в значительной степени игнорировал его в течение нескольких десятилетий 1 . До появления графических пользовательских интерфейсов 2 , которые, как оказалось, действительно нуждаются в ООП. Когда вы нажимаете кнопку в окне, что может быть проще для генерации соответствующих ответов, чем отправить этой кнопке (или ее суррогату) сообщение Click 3 ?
После этого полетел ООП. Было написано множество 4 книг и выпущено бесчисленное количество 5 статей. Итак, теперь все в деталях разбираются в объектно-ориентированном программировании, верно?
Итак, теперь все в деталях разбираются в объектно-ориентированном программировании, верно?
К сожалению, код (и Интернет) говорят нет.
Самым большим вопросом путаницы и разногласий кажется композиция против наследования, часто резюмируемая мантрой «отдавайте предпочтение композиции наследованию». Давайте поговорим об этом.
Мантры считаются вредными
Как эвристика, «предпочесть композицию наследованию» — это нормально, однако я не фанат мантр. Хотя они часто содержат зерно истины, люди слишком легко слышат лозунг, не понимая его источника или контекста, и, таким образом, избегают думать самостоятельно, а это никогда не заканчивается хорошо.
Я также не являюсь поклонником нелепых кликбейтных заголовков, таких как «Наследование — это зло» 6 , особенно когда автор пытается подкрепить такое возмутительное утверждение ненадлежащим использованием наследования… а затем обвиняет его. Как плотник, заявляющий, что молотки бесполезны, потому что они плохо закручивают шурупы.
Начнем с основ.
Определения
Вот определение объектно-ориентированного программирования, которое я буду использовать в оставшейся части статьи: предположим, что у нас есть «классический» язык ООП, то есть такой, который поддерживает классы с полями, методами и одиночным наследованием. Никаких интерфейсов, никаких примесей, никаких аспектов, никакого множественного наследования, никаких делегатов, никаких замыканий, никаких лямбда-выражений, ничего, кроме основ:
- Класс: именованное понятие в доменном пространстве с необязательным суперклассом, определенным как набор полей и методов.
- Поле: именованное свойство некоторого типа, которое может ссылаться на другой объект (см. состав)
- Метод: именованная функция или процедура с параметрами или без них, реализующая некоторое поведение класса.
- Наследование: класс может наследовать — использовать по умолчанию — поля и методы своего суперкласса. Наследование является транзитивным, поэтому класс может наследовать от другого класса, который наследуется от другого класса, и так далее, вплоть до базового класса (обычно Object, возможно, неявного/отсутствующего). Подклассы могут переопределять некоторые методы и/или поля, чтобы изменить поведение по умолчанию.
- Состав: когда тип поля является классом, поле будет содержать ссылку на другой объект, тем самым создавая ассоциативное отношение между ними. Не вдаваясь в нюансы разницы между простой ассоциацией, агрегацией и композицией, давайте интуитивно определим композицию как случай, когда класс использует другой объект для обеспечения части или всей своей функциональности.
- Инкапсуляция: взаимодействуя с объектами, а не напрямую с реализацией методов и полей, мы скрываем и защищаем реализацию класса. Если потребитель ничего не знает об объекте, кроме его открытого интерфейса, он не может полагаться на какие-либо детали внутренней реализации.
 Наследование является транзитивным, поэтому класс может наследовать от другого класса, который наследуется от другого класса, и так далее, вплоть до базового класса (обычно Object, возможно, неявного/отсутствующего). Подклассы могут переопределять некоторые методы и/или поля, чтобы изменить поведение по умолчанию.
Наследование является транзитивным, поэтому класс может наследовать от другого класса, который наследуется от другого класса, и так далее, вплоть до базового класса (обычно Object, возможно, неявного/отсутствующего). Подклассы могут переопределять некоторые методы и/или поля, чтобы изменить поведение по умолчанию.Наследование является основным
Наследование является фундаментальным для объектно-ориентированного программирования. Язык программирования может иметь объекты и сообщения, но без наследования он не является объектно-ориентированным (просто «объектно-ориентированным», но все же полиморфным).
Язык программирования может иметь объекты и сообщения, но без наследования он не является объектно-ориентированным (просто «объектно-ориентированным», но все же полиморфным).
…а также Состав
Композиция также является фундаментальной для каждого языка . Даже если язык не поддерживает композицию (редкость в наши дни!), люди все еще думают в терминах частей и компонентов. Было бы невозможно разбить сложные проблемы на модульные решения без композиции.
(Инкапсуляция тоже фундаментальна, но мы не будем здесь подробно о ней говорить.)
Итак, о чем спор?
И композиция, и наследование фундаментальны, так в чем же дело?
Большое дело в том, чтобы думать, что одно может заменить другое во всех случаях, или что одно лучше или хуже другого. Как и во всем остальном в разработке программного обеспечения, здесь приходится идти на компромиссы.
Композицию довольно легко понять — мы можем видеть композицию в повседневной жизни: у стула есть ножки, стена состоит из кирпичей и раствора и так далее. Хотя определение наследования простое, оно может стать сложной и запутанной вещью при неразумном использовании. Наследование — это скорее абстракция, о которой мы можем только говорить, а не касаться напрямую. Хотя во многих ситуациях можно имитировать наследование с помощью композиции, часто это оказывается громоздким. Цель композиции очевидна: сделать из частей целое. Цель наследования немного сложнее, потому что наследование служит двум целям: семантике и механике.
Хотя определение наследования простое, оно может стать сложной и запутанной вещью при неразумном использовании. Наследование — это скорее абстракция, о которой мы можем только говорить, а не касаться напрямую. Хотя во многих ситуациях можно имитировать наследование с помощью композиции, часто это оказывается громоздким. Цель композиции очевидна: сделать из частей целое. Цель наследования немного сложнее, потому что наследование служит двум целям: семантике и механике.
Семантика наследования
Наследование фиксирует семантику (значение) в иерархии классификации (таксономии), упорядочивая понятия от обобщенных до специализированных, группируя связанные понятия в поддеревья и т.д. Семантика класса в основном определяется его интерфейсом, набором сообщений, на которые он отвечает, но часть семантики также содержится в наборе сообщений, которые отправляет класс. При наследовании от класса вы неявно принимаете на себя ответственность за все сообщения, которые суперкласс отправляет от вашего имени, а не только за сообщения, которые он может получить. Это делает подкласс более тесно связанным со своим суперклассом, чем если бы он просто использовал экземпляр суперкласса в качестве компонента, а не наследовал от него. Обратите внимание, что даже в классах, которые мало что «делают», имя класса сообщает разработчику важную семантическую информацию о предметной области.
Это делает подкласс более тесно связанным со своим суперклассом, чем если бы он просто использовал экземпляр суперкласса в качестве компонента, а не наследовал от него. Обратите внимание, что даже в классах, которые мало что «делают», имя класса сообщает разработчику важную семантическую информацию о предметной области.
Механика наследования
Наследование захватывает механику, кодируя представление данных (полей) и поведения (методов) класса и делая его доступным для повторного использования и расширения в подклассах. Механически подкласс наследует реализацию суперкласса и, следовательно, его интерфейс.
Двойная цель наследования 7 в большинстве современных ООП-языков, как мне кажется, является причиной большей путаницы. Многие думают, что «повторное использование кода» — основная цель наследования, но это не единственная его цель. Чрезмерный акцент на повторном использовании может привести к трагически ошибочным проектам. Давайте посмотрим на пару примеров.
Давайте посмотрим на пару примеров.
Как неправильно использовать наследование — пример 1
Начнем с простого и чрезвычайно распространенного примера неправильного использования наследования:
class Stack extends ArrayList {
public void push(значение объекта) {…}
общедоступный объект pop() { … }
}
Этот класс будет функционировать как стек, но его интерфейс сильно раздут. Открытый интерфейс этого класса не только push и pop, как можно было бы ожидать от класса с именем Stack, он также включает в себя получение, установку, добавление, удаление, очистку и кучу других сообщений, унаследованных от ArrayList, которые не подходят для класса Stack. Куча.
Вы можете переопределить все нежелательные методы и, возможно, адаптировать некоторые полезные (например, очистку), но для того, чтобы скрыть ошибку моделирования, потребуется много работы. На самом деле три ошибки моделирования, одна семантическая, одна механическая, одна и та, и другая:
- Семантически утверждение «Стек — это ArrayList» неверно; Stack не является правильным подтипом ArrayList. Предполагается, что стек обеспечивает соблюдение принципа «последним пришел — первым вышел» — ограничение, которое легко удовлетворяется интерфейсом push/pop, но не обеспечивается интерфейсом ArrayList.
- Механически наследование от ArrayList нарушает инкапсуляцию; использование ArrayList для хранения коллекции объектов стека — это выбор реализации, который должен быть скрыт от потребителей.
- Наконец, реализация стека путем наследования от ArrayList представляет собой междоменное отношение: ArrayList — коллекция с произвольным доступом; Стек — это концепция организации очереди со специально ограниченным (неслучайным) доступом 8 . Это разные области моделирования.
 Предполагается, что стек обеспечивает соблюдение принципа «последним пришел — первым вышел» — ограничение, которое легко удовлетворяется интерфейсом push/pop, но не обеспечивается интерфейсом ArrayList.
Предполагается, что стек обеспечивает соблюдение принципа «последним пришел — первым вышел» — ограничение, которое легко удовлетворяется интерфейсом push/pop, но не обеспечивается интерфейсом ArrayList.Последняя проблема важна, но немного тонка, поэтому давайте рассмотрим ее на другом примере.
Как неправильно использовать наследование — пример 2
Создание класса предметной области путем наследования от класса реализации является распространенным злоупотреблением наследованием. Например, предположим, что мы хотим что-то сделать с определенным сегментом наших клиентов. Самый простой и очевидный способ — создать подкласс ArrayList
Например, предположим, что мы хотим что-то сделать с определенным сегментом наших клиентов. Самый простой и очевидный способ — создать подкласс ArrayList
Неправильно. Это будут отношения междоменного наследования, и их следует избегать:
- ArrayList
уже является подклассом списка, набор утилит — класс реализации . - CustomerGroup — это еще один подкласс — класс домена .
- Классы предметной области должны использовать классы реализации , а не наследовать от них.
Пространство реализации должно быть невидимым на уровне домена. Когда мы думаем о том, что делает наше программное обеспечение, мы работаем на уровне предметной области; мы не хотим отвлекаться на детали того, как он работает. Если мы сосредоточимся только на «повторном использовании кода» через наследование, мы будем постоянно попадать в эту ловушку.
Одиночное наследование не является проблемой
Одиночное наследование по-прежнему является наиболее распространенной моделью ООП; одиночное наследование обязательно является наследованием реализации, что может привести к сильной связи между классами. Проблема, по-видимому, заключается в том, что у нас есть только один путь наследования, который можно использовать для моделирования как наших механических, так и семантических потребностей. Если вы используете его для одного, вы не можете использовать его для другого. Так не решает ли множественное наследование эту проблему?
№ . Отношения наследования не должны пересекать границы домена (домен реализации и домен приложения). Наследование CustomerGroup от ArrayList
Предпочтительное (по крайней мере для меня!) решение состоит в том, чтобы наследовать от служебных классов столько, сколько необходимо для реализации ваших механических структур, а затем использовать эти структуры в классах предметной области посредством композиции, а не наследования. Позвольте мне повторить это:
Позвольте мне повторить это:
Если вы не создаете класс реализации, вы не должны наследовать от класса реализации.
Это одна из самых распространенных проблем новичков — потому что это так удобно! — а причины, по которым это неправильно, нечасто обсуждаются в литературе по программированию, поэтому я повторю еще раз: ваши классы предметной области должны использовать классы реализации , а не быть одним из них. Разделяйте эти таксономии/домены.
Итак, когда и как мы должны использовать наследование?
Использование скважины наследства
Наиболее распространенное и выгодное использование наследования для дифференциального программирования . Нам нужен виджет, похожий на существующий класс Widget, но с некоторыми изменениями и улучшениями. В этом случае наследуйте; это уместно, потому что наш подкласс по-прежнему является виджетом, мы хотим повторно использовать весь интерфейс и реализацию из суперкласса, и наши изменения в основном добавка . Если вы обнаружите, что ваш подкласс удаляет вещи, предоставленные суперклассом, задайте вопрос о наследовании от этого суперкласса.
Если вы обнаружите, что ваш подкласс удаляет вещи, предоставленные суперклассом, задайте вопрос о наследовании от этого суперкласса.
Наследование наиболее полезно для группировки связанных наборов понятий, идентификации семейств классов и в целом для организации имен и понятий, описывающих предметную область. По мере того, как мы углубляемся в реализацию системы, мы можем обнаружить, что наши первоначальные обобщения концепций предметной области, зафиксированные в наших иерархиях наследования, начинают разрушаться. Не бойтесь разбирать иерархии наследования на наборы взаимодополняющих взаимодействующих интерфейсов и компонентов, когда код ведет вас в этом направлении 9 .
Как решить: состав или наследство?
Если у вас есть ситуация, когда сработает либо композиция, либо наследование, рассмотрите возможность разделения обсуждения дизайна на две части:
- Представление/реализация концепций вашей предметной области является одним измерением
- Семантика понятий вашей предметной области и их связь друг с другом — это второе измерение
В общем, наследование в пределах одного из этих измерений — это нормально. Проблема возникает, когда мы забываем разделить два измерения и начинаем наследовать через межпространственные границы.
Проблема возникает, когда мы забываем разделить два измерения и начинаем наследовать через межпространственные границы.
Если вы обнаружите, что используете компонент для обеспечения подавляющего большинства своих функций, создавая методы переадресации в своем классе для вызова методов компонента, показывая поля компонента и т. д., рассмотрите возможность наследования — для некоторых или всех функций. желаемое поведение — может быть более подходящим.
Нет никакой замены объектному моделированию и критическому проектному мышлению. Но если вам нужны какие-то рекомендации, рассмотрите их —
Наследование следует использовать только в следующих случаях:
- Оба класса находятся в одном логическом домене
- Подкласс является правильным подтипом надкласса
- Реализация суперкласса необходима или подходит для подкласса
- Улучшения, сделанные подклассом, в основном аддитивны.
Бывают моменты, когда все эти вещи сходятся:
- Моделирование предметной области более высокого уровня
- Фреймворки и расширения фреймворков
- Дифференциальное программирование
Если вы не делаете ни одной из этих вещей, вам, вероятно, не понадобится очень часто наследование классов. «Предпочтение» композиции — это не вопрос «лучше», это вопрос «наиболее подходящего» для ваших нужд в конкретном контексте.
«Предпочтение» композиции — это не вопрос «лучше», это вопрос «наиболее подходящего» для ваших нужд в конкретном контексте.
Надеемся, что эти рекомендации помогут вам заметить разницу.
Удачного кодирования!
Приложение
Особая благодарность следующим мыслителям за ценный вклад и комментарии: Питу Ходжсону, Тиму Брауну, Скотту Робинсону, Мартину Фаулеру, Минди Ор, Шону Ньюэму, Сэму Гибсону и Махендре Кария.
1. Первый официально объектно-ориентированный язык SIMULA 67 родился в 1967 году. Объектно-ориентированному программированию исполнилось 48 лет!
2. Программисты систем и приложений перешли на C++ в середине 1980-х, но повсеместное распространение ООП пришлось ждать еще десятилетие.
3. да, я упрощаю, игнорируя слушателей/делегатов событий/и т.д.; стараясь сделать эту статью короткой!
4. На момент написания этой статьи Amazon заявляет о 24 777 книгах по теме объектно-ориентированного программирования
5.